

Evolution of regulatory signatures in primate cortical neurons at cell-type resolution
- PMID: 33109720
- PMCID: PMC7668098
- DOI: 10.1073/pnas.2011884117
Evolution of regulatory signatures in primate cortical neurons at cell-type resolution
Abstract
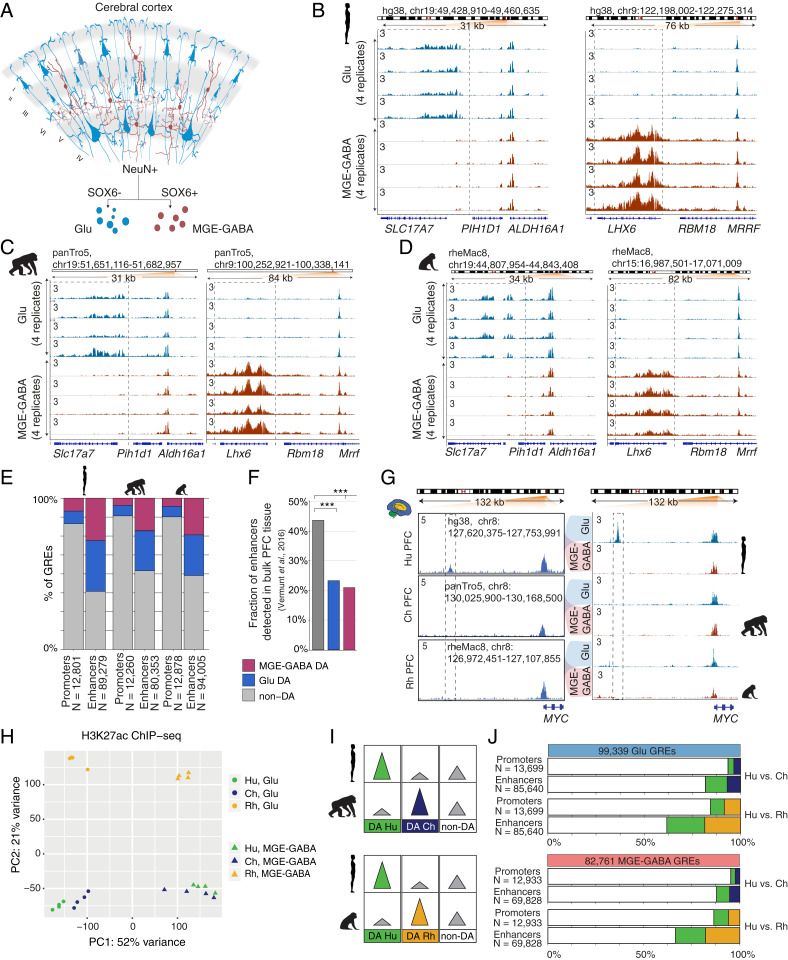
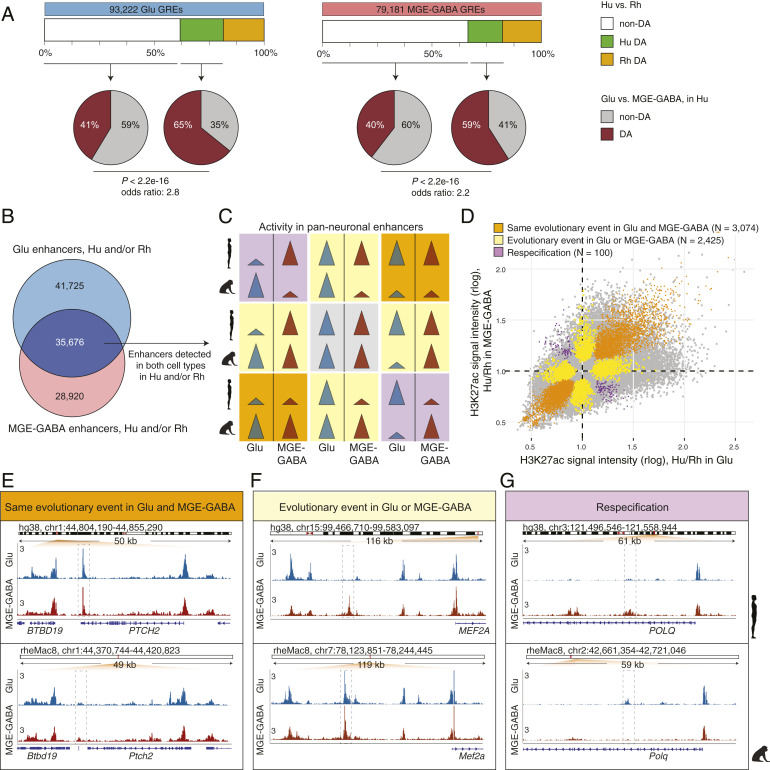
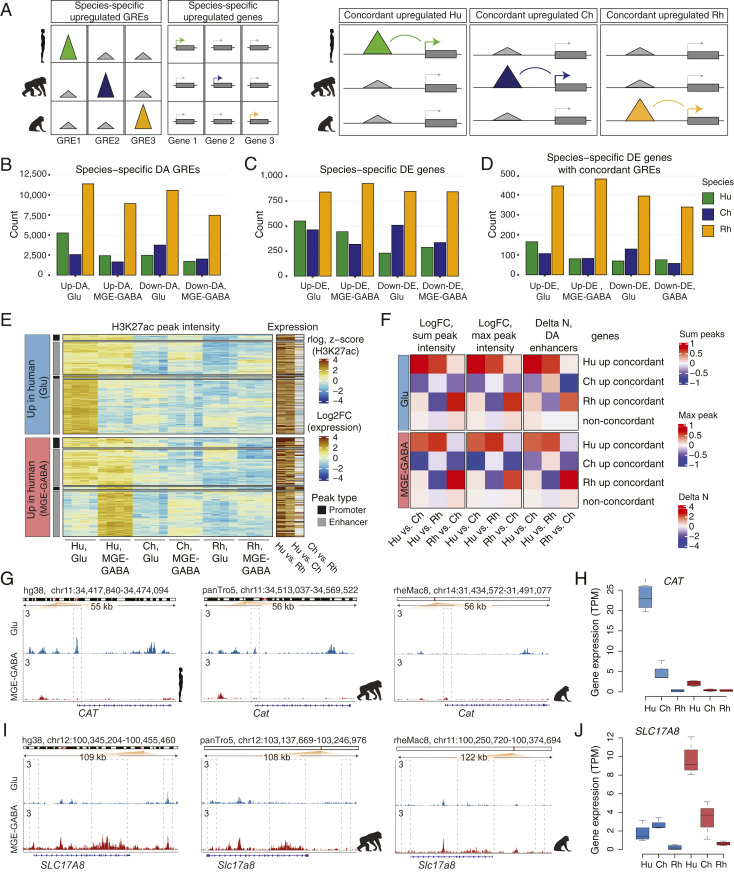
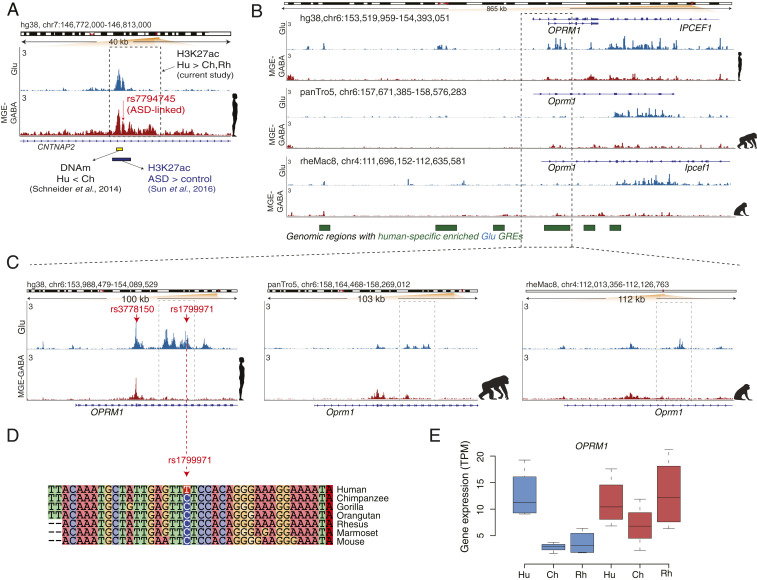
The human cerebral cortex contains many cell types that likely underwent independent functional changes during evolution. However, cell-type-specific regulatory landscapes in the cortex remain largely unexplored. Here we report epigenomic and transcriptomic analyses of the two main cortical neuronal subtypes, glutamatergic projection neurons and GABAergic interneurons, in human, chimpanzee, and rhesus macaque. Using genome-wide profiling of the H3K27ac histone modification, we identify neuron-subtype-specific regulatory elements that previously went undetected in bulk brain tissue samples. Human-specific regulatory changes are uncovered in multiple genes, including those associated with language, autism spectrum disorder, and drug addiction. We observe preferential evolutionary divergence in neuron subtype-specific regulatory elements and show that a substantial fraction of pan-neuronal regulatory elements undergoes subtype-specific evolutionary changes. This study sheds light on the interplay between regulatory evolution and cell-type-dependent gene-expression programs, and provides a resource for further exploration of human brain evolution and function.
Keywords: GABAergic neurons; H3K27ac histone modification; glutamatergic neurons; primate evolution; regulatory elements.
Copyright © 2020 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Ma T. et al., Subcortical origins of human and monkey neocortical interneurons. Nat. Neurosci. 16, 1588–1597 (2013). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
