

Biotic Interactions Are More Important than Propagule Pressure in Microbial Community Invasions
- PMID: 33109758
- PMCID: PMC7593967
- DOI: 10.1128/mBio.02089-20
Biotic Interactions Are More Important than Propagule Pressure in Microbial Community Invasions
Abstract
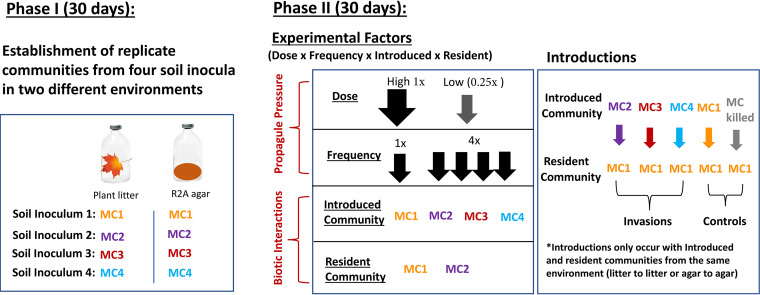
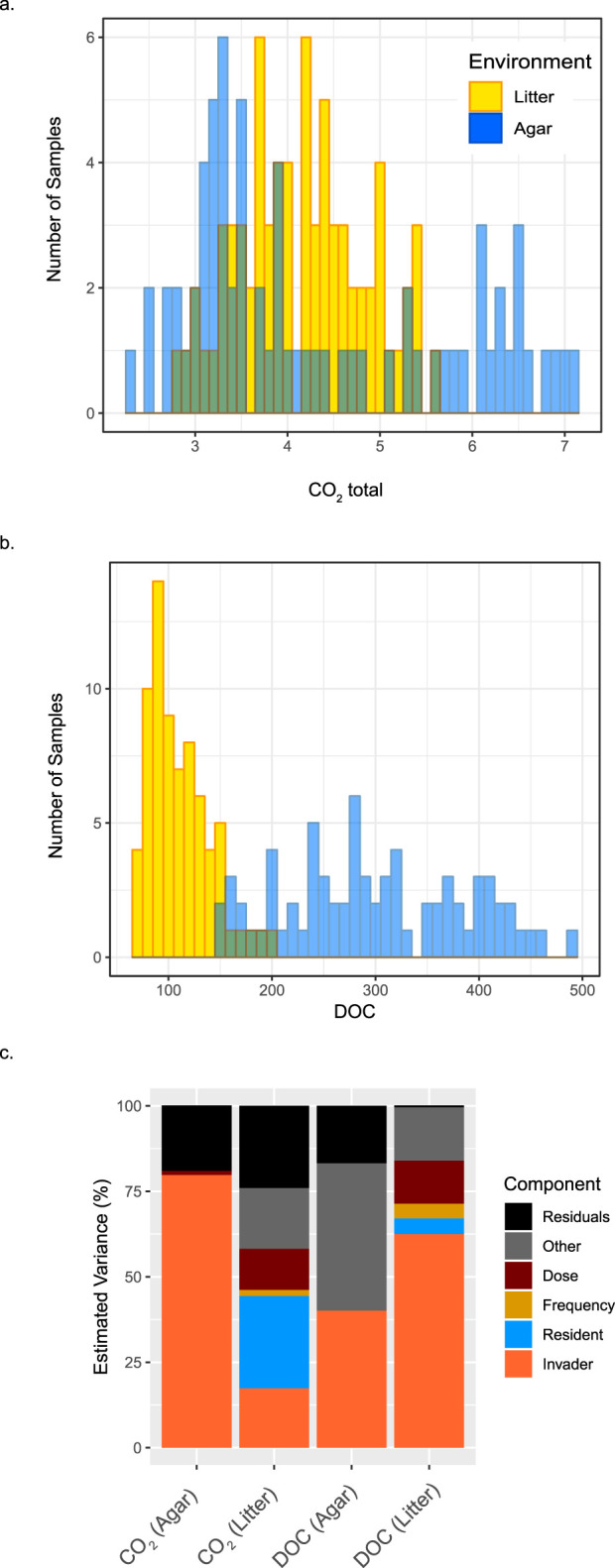
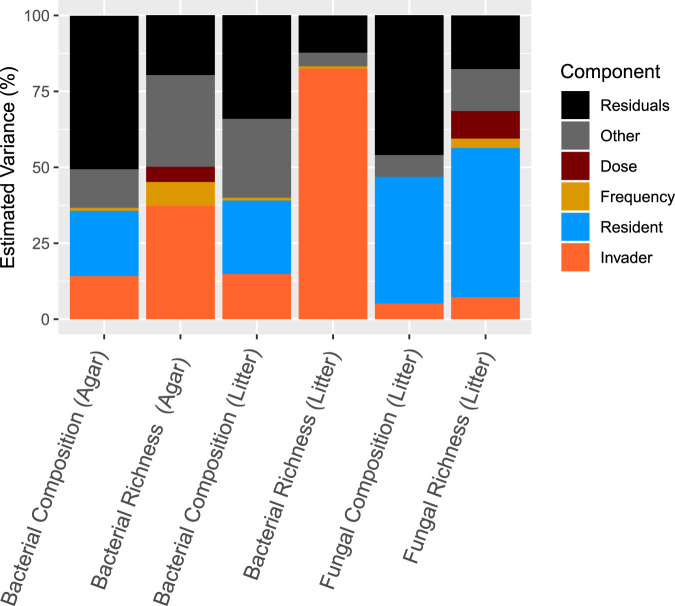
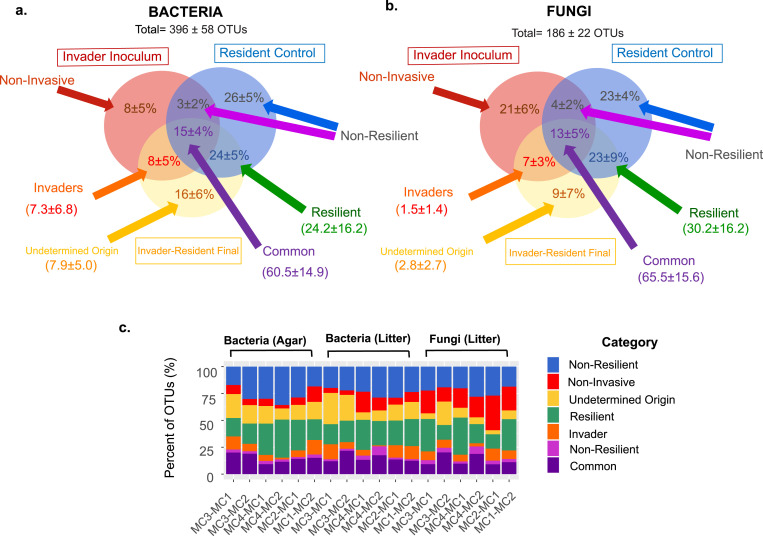
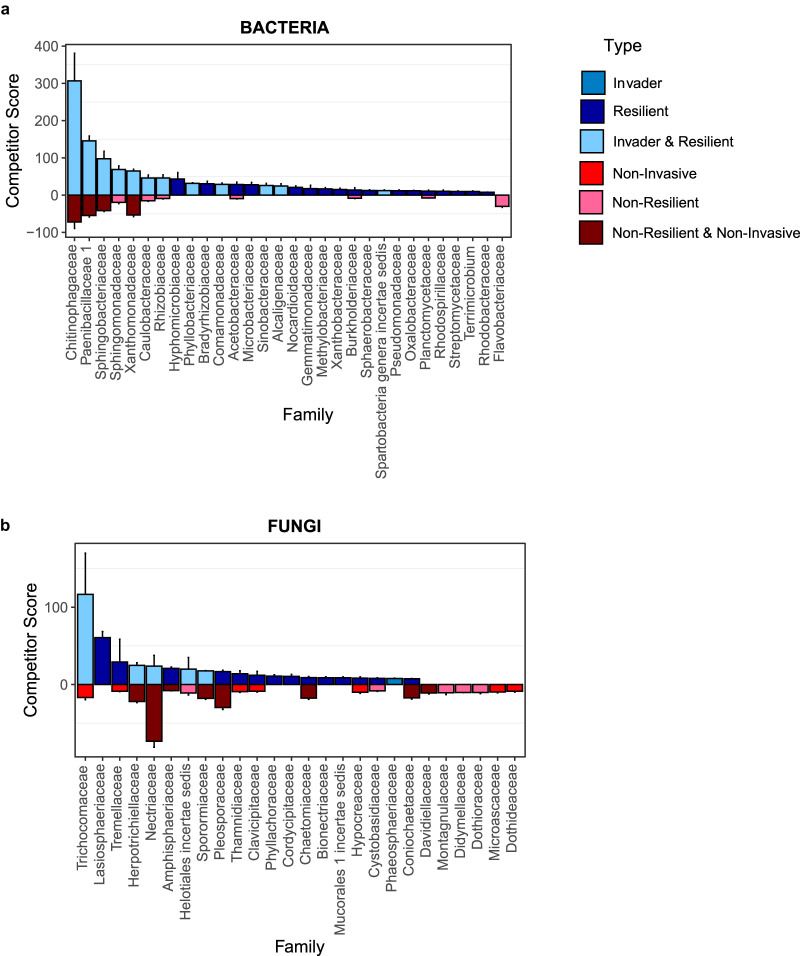
Microbial probiotics are intended to improve functions in diverse ecosystems, yet probiotics often fail to establish in a preexisting microbiome. This is a species invasion problem. The relative importance of the two major factors controlling establishment in this context-propagule pressure (inoculation dose and frequency) and biotic interactions (composition of introduced and resident communities)-is unknown. We tested the effect of these factors in driving microbial composition and functioning following 12 microbial community invasions (e.g., introductions of many microbial invaders) in microcosms. Ecosystem functioning over a 30-day postinvasion period was assessed by measuring activity (respiration) and environment modification (dissolved organic carbon abundance). To test the dependence on environmental context, experiments were performed in two resource environments. In both environments, biotic interactions were more important than propagule pressure in driving microbial composition and community function, but the magnitude of effect varied by environment. Successful invaders comprised approximately 8% of the total number of operational taxonomic units (OTUs). Bacteria were better invaders than fungi, with average relative abundances of 7.4% ± 6.8% and 1.5% ± 1.4% of OTUs, respectively. Common bacterial invaders were associated with stress response traits. The most resilient bacterial and fungal families, in other words, those least impacted by invasions, were linked to antimicrobial resistance or production traits. Illuminating the principles that determine community composition and functioning following microbial invasions is key to efficient community engineering.IMPORTANCE With increasing frequency, humans are introducing new microbes into preexisting microbiomes to alter functioning. Example applications include modification of microflora in human guts for better health and those of soil for food security and/or climate management. Probiotic applications are often approached as trial-and-error endeavors and have mixed outcomes. We propose that increased success in microbiome engineering may be achieved with a better understanding of microbial invasions. We conducted a microbial community invasion experiment to test the relative importance of propagule pressure and biotic interactions in driving microbial community composition and ecosystem functioning in microcosms. We found that biotic interactions were more important than propagule pressure in determining the impact of microbial invasions. Furthermore, the principles for community engineering vary among organismal groups (bacteria versus fungi).
Keywords: bacterial traits; ecosystem functioning; ecosystem manipulation; fungal traits; invasion biology; microbial composition; microbiome engineering; probiotics.
Copyright © 2020 Albright et al.
Figures
References
-
- Bakken JS, Borody T, Brandt LJ, Brill JV, Demarco DC, Franzos MA, Kelly C, Khoruts A, Louie T, Martinelli LP, Moore TA, Russell G, Surawicz C, Fecal Microbiota Transplantation Workgroup. 2011. Treating Clostridium difficile infection with fecal microbiota transplantation. Clin Gastroenterol Hepatol 9:1044–1049. doi: 10.1016/j.cgh.2011.08.014. - DOI - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
