

Viral tools for neuroscience
- PMID: 33110222
- PMCID: PMC7808553
- DOI: 10.1038/s41583-020-00382-z
Viral tools for neuroscience
Abstract
Recombinant viruses are the workhorse of modern neuroscience. Whether one would like to understand a neuron's morphology, natural activity patterns, molecular composition, connectivity or behavioural and physiologic function, most studies begin with the injection of an engineered virus, often an adeno-associated virus or herpes simplex virus, among many other types. Recombinant viruses currently enable some combination of cell type-specific, circuit-selective, activity-dependent and spatiotemporally resolved transgene expression. Viruses are now used routinely to study the molecular and cellular functions of a gene within an identified cell type in the brain, and enable the application of optogenetics, chemogenetics, calcium imaging and related approaches. These advantageous properties of engineered viruses thus enable characterization of neuronal function at unprecedented resolution. However, each virus has specific advantages and disadvantages, which makes viral tool selection paramount for properly designing and executing experiments within the central nervous system. In the current Review, we discuss the key principles and uses of engineered viruses and highlight innovations that are needed moving forward.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
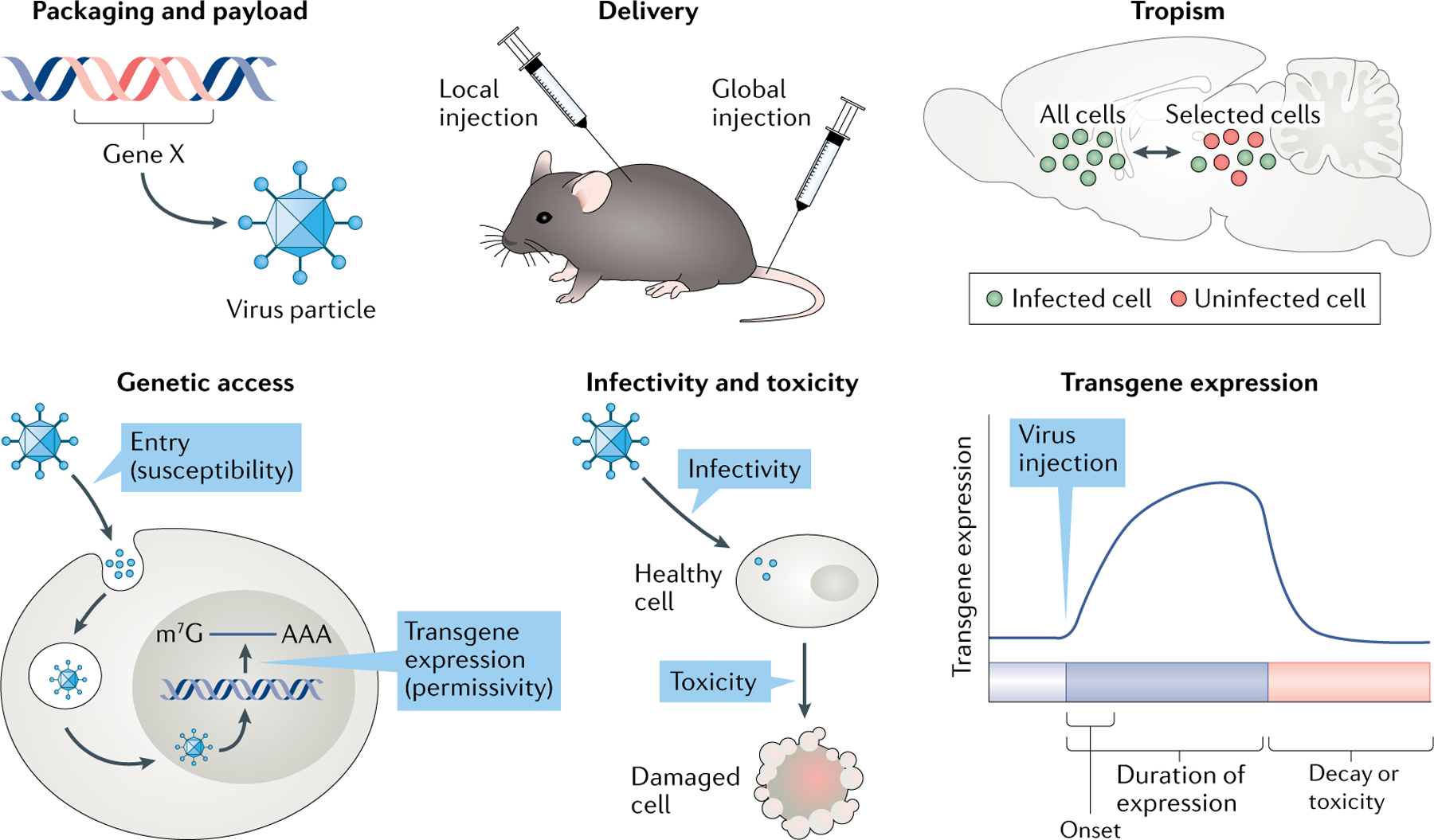
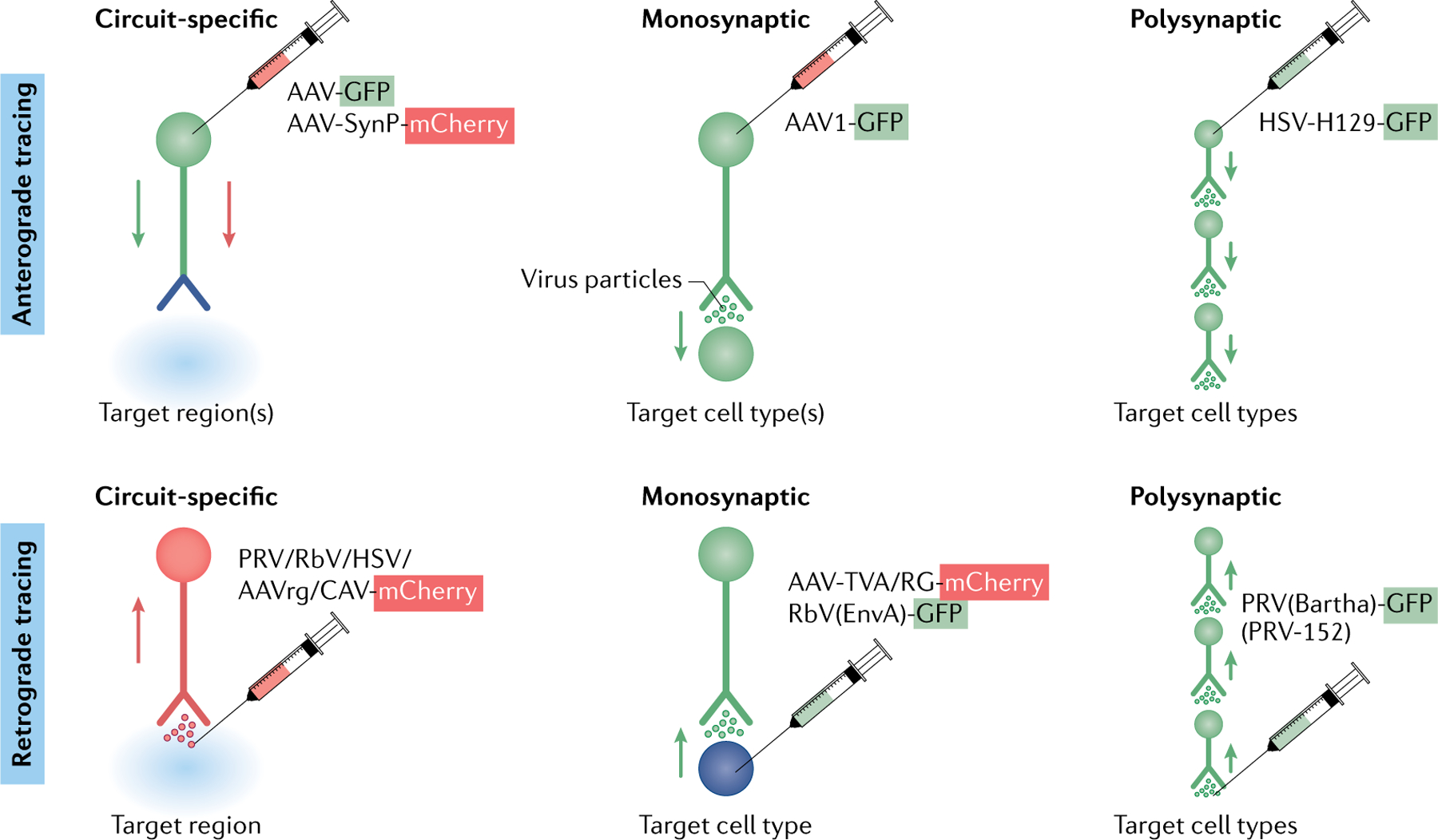
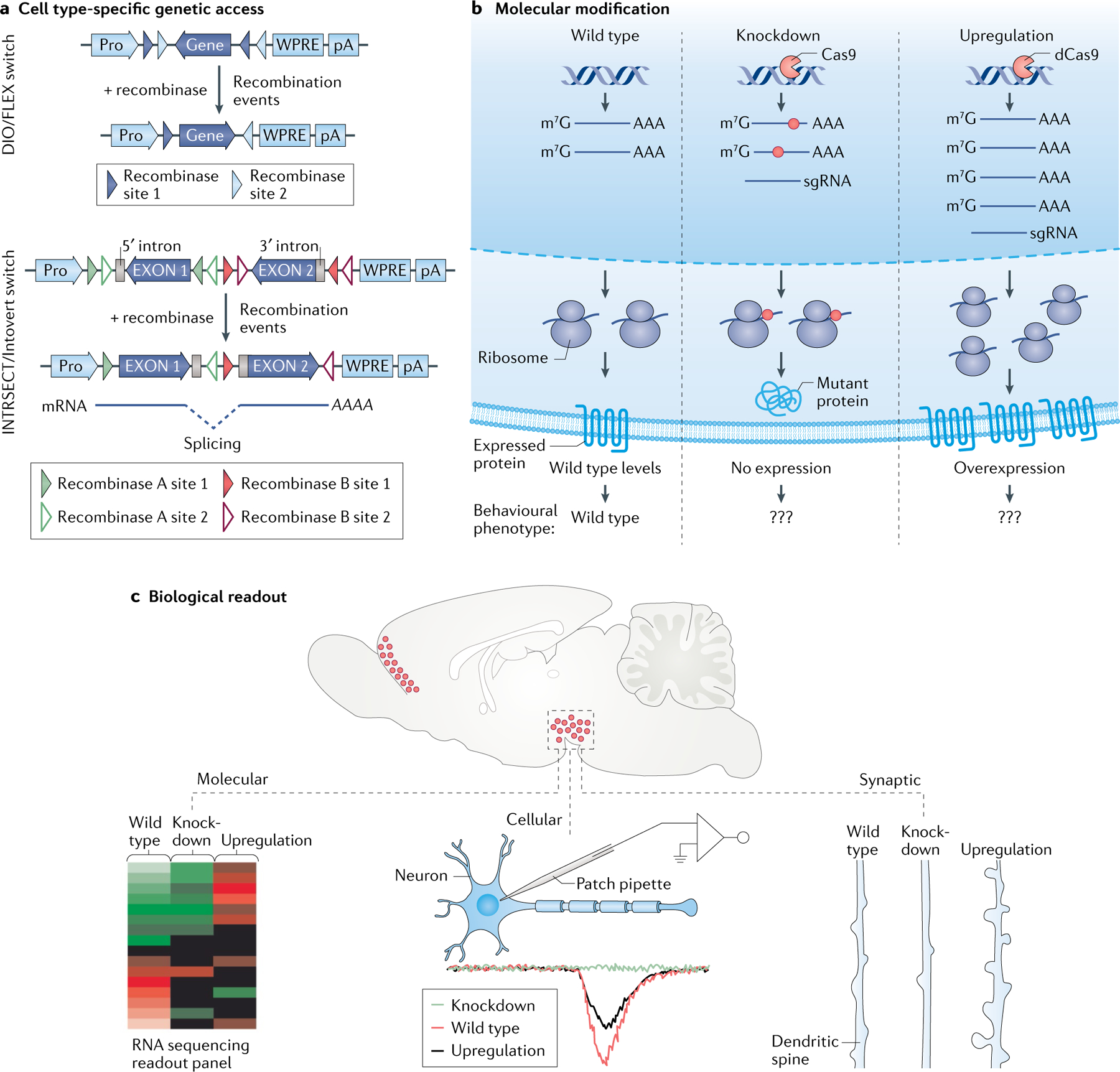
References
-
- Flint SJ, Racaniello VR, Rall GG, Skalka AM & Enquist LW Principles of Virology 4th edn Vol. 4 (ASM, 2015).
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
