

TET2-Loss-of-Function-Driven Clonal Hematopoiesis Exacerbates Experimental Insulin Resistance in Aging and Obesity
- PMID: 33113366
- PMCID: PMC7856871
- DOI: 10.1016/j.celrep.2020.108326
TET2-Loss-of-Function-Driven Clonal Hematopoiesis Exacerbates Experimental Insulin Resistance in Aging and Obesity
Abstract
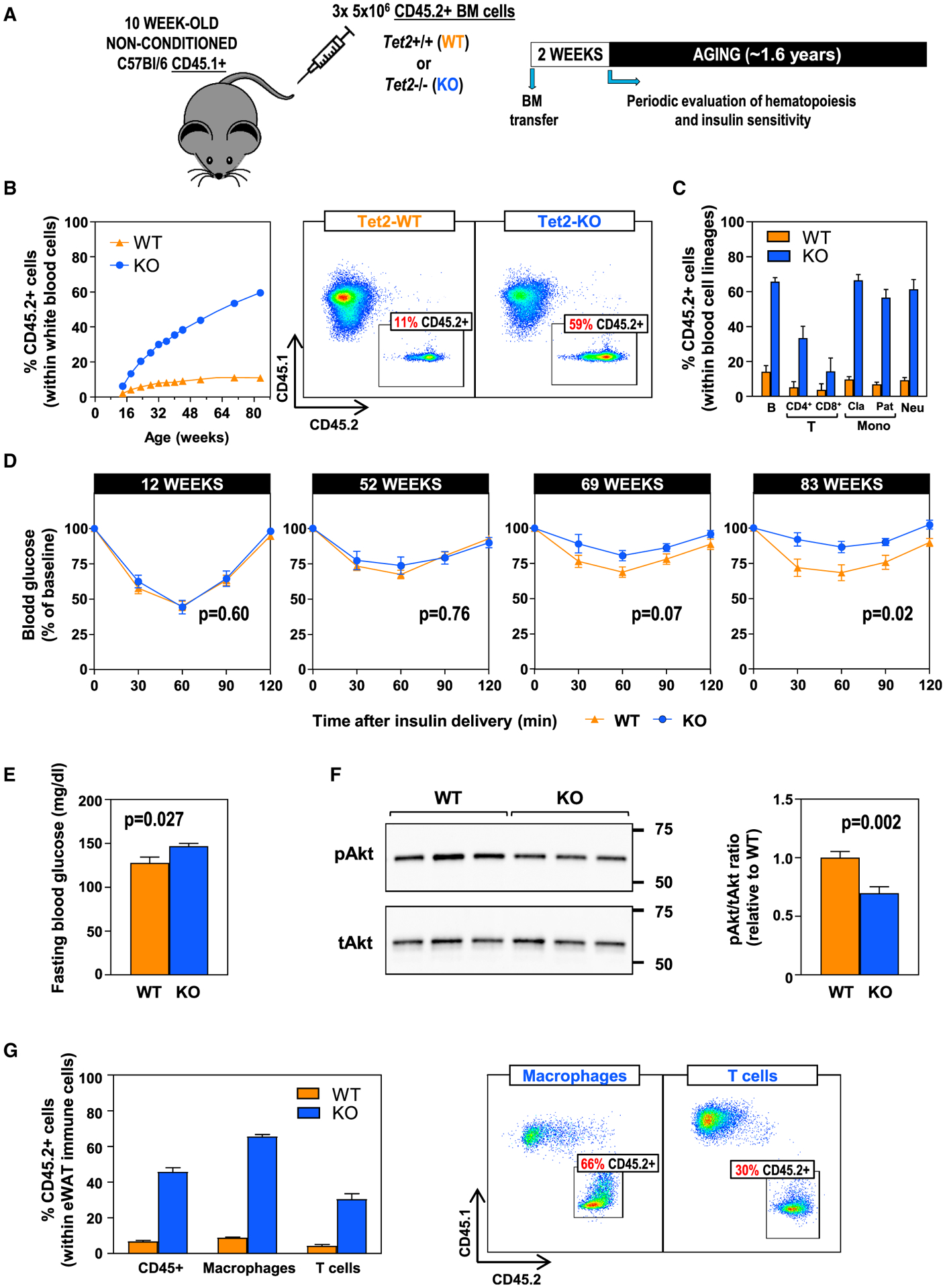
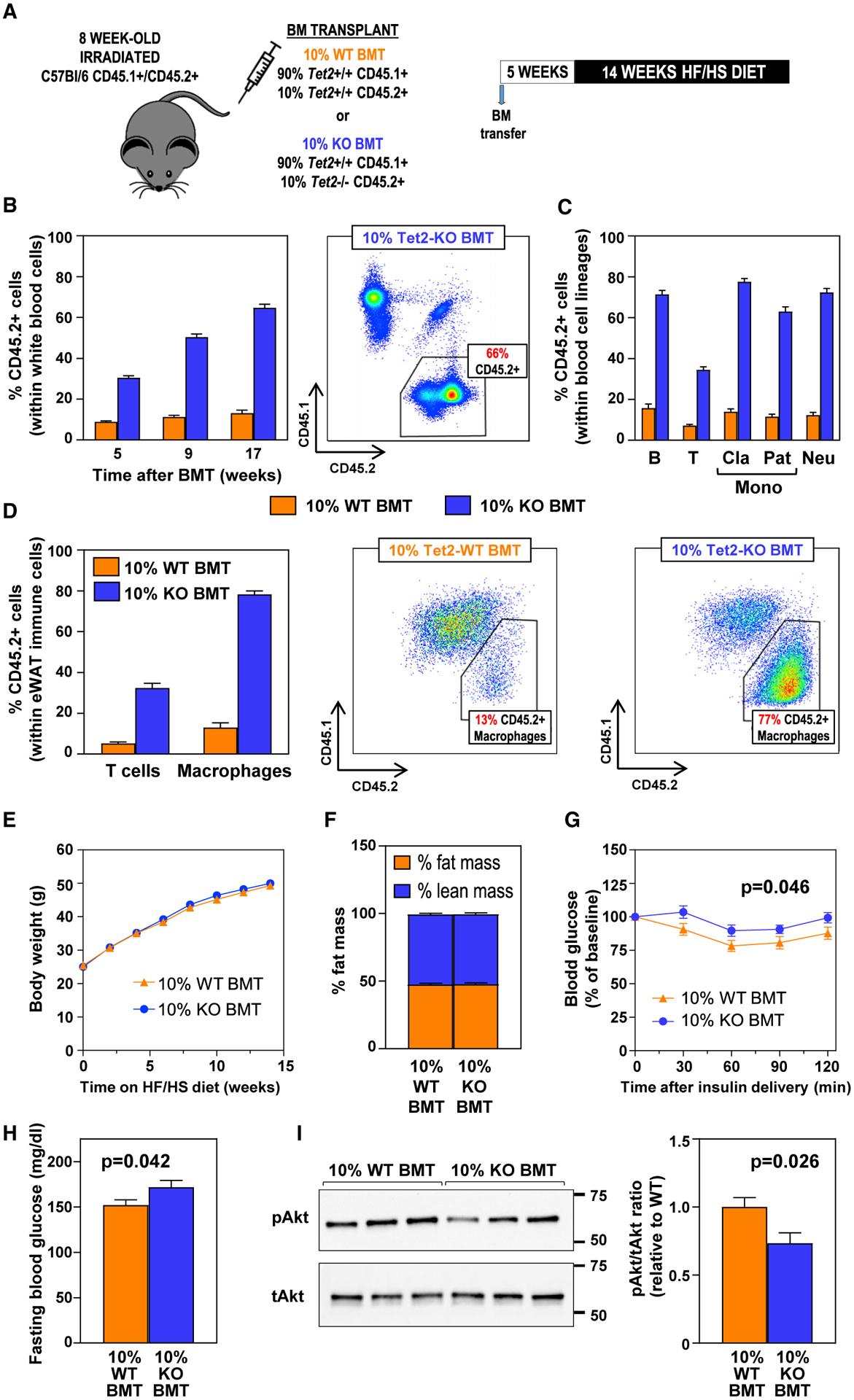
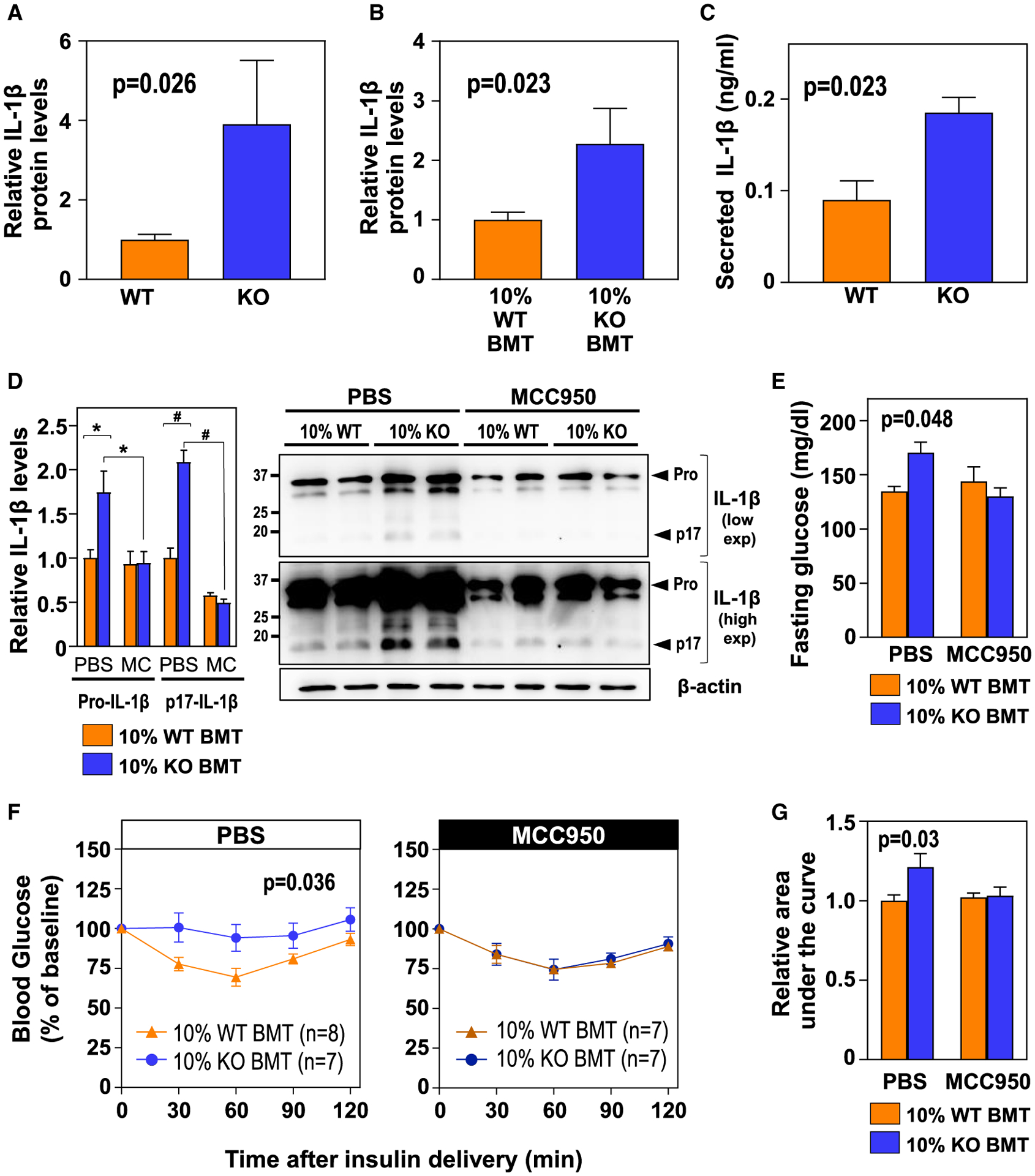
Human aging is frequently accompanied by the acquisition of somatic mutations in the hematopoietic system that induce clonal hematopoiesis, leading to the development of a mutant clone of hematopoietic progenitors and leukocytes. This somatic-mutation-driven clonal hematopoiesis has been associated with an increased incidence of cardiovascular disease and type 2 diabetes, but whether this epidemiological association reflects a direct, causal contribution of mutant hematopoietic and immune cells to age-related metabolic abnormalities remains unexplored. Here, we show that inactivating mutations in the epigenetic regulator TET2, which lead to clonal hematopoiesis, aggravate age- and obesity-related insulin resistance in mice. This metabolic dysfunction is paralleled by increased expression of the pro-inflammatory cytokine IL-1β in white adipose tissue, and it is suppressed by pharmacological inhibition of NLRP3 inflammasome-mediated IL-1β production. These findings support a causal contribution of somatic TET2 mutations to insulin resistance and type 2 diabetes.
Keywords: CHIP; IL-1β; TET2; adipose tissue; aging; clonal hematopoiesis; diabetes; insulin resistance; obesity; somatic mutations.
Copyright © 2020 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests K.W. and J.J.F. are co-inventors on a patent related to the treatment of cardiometabolic diseases associated with somatic TET2 mutations. M.A.C. is co-founder and chief executive officer of Inflazome, a company developing drugs that target inflammasomes. These authors have no additional conflicts of interest. Other authors declare no competing interests.
Figures
References
-
- Abplanalp WT, Mas-Peiro S, Cremer S, John D, Dimmeler S, and Zeiher AM (2020). Association of Clonal Hematopoiesis of Indeterminate Potential With Inflammatory Gene Expression in Patients With Severe Degenerative Aortic Valve Stenosis or Chronic Postischemic Heart Failure. JAMA Cardiol. Published online July 8, 2020. In press. 10.1001/jamacardio.2020.2468. - DOI - PMC - PubMed
-
- Arends CM, Galan-Sousa J, Hoyer K, Chan W, Jäger M, Yoshida K, Seemann R, Noerenberg D, Waldhueter N, Fleischer-Notter H, et al. (2018). Hematopoietic lineage distribution and evolutionary dynamics of clonal hematopoiesis. Leukemia 32, 1908–1919. - PubMed
-
- Bick AG, Weinstock JS, Nandakumar SK, Fulco CP, Leventhal MJ, Bao EL, Nasser J, Zekavat SM, Szeto MD, Laurie C, et al. (2019). Inherited Causes of Clonal Hematopoiesis of Indeterminate Potential in TOPMed Whole Genomes. bioRxiv. 10.1101/782748. - DOI
-
- Bonnefond A, Skrobek B, Lobbens S, Eury E, Thuillier D, Cauchi S, Lantieri O, Balkau B, Riboli E, Marre M, et al. (2013). Association between large detectable clonal mosaicism and type 2 diabetes with vascular complications. Nat. Genet 45, 1040–1043. - PubMed
-
- Buscarlet M, Provost S, Zada YF, Barhdadi A, Bourgoin V, Lépine G, Mollica L, Szuber N, Dubé MP, and Busque L (2017). DNMT3A and TET2 dominate clonal hematopoiesis and demonstrate benign phenotypes and different genetic predispositions. Blood 130, 753–762. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
