

Strategies for cystic fibrosis transmembrane conductance regulator inhibition: from molecular mechanisms to treatment for secretory diarrhoeas
- PMID: 33113586
- PMCID: PMC7756540
- DOI: 10.1002/1873-3468.13971
Strategies for cystic fibrosis transmembrane conductance regulator inhibition: from molecular mechanisms to treatment for secretory diarrhoeas
Abstract
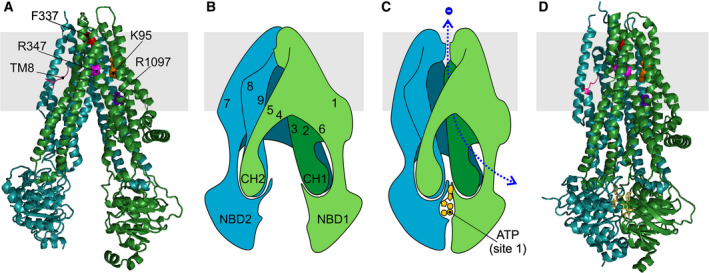
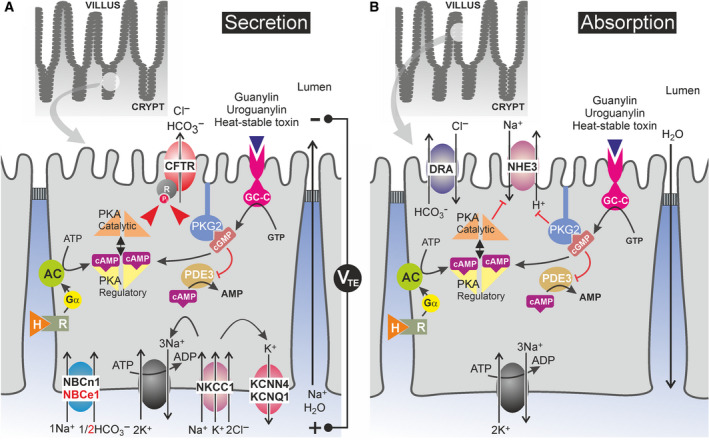
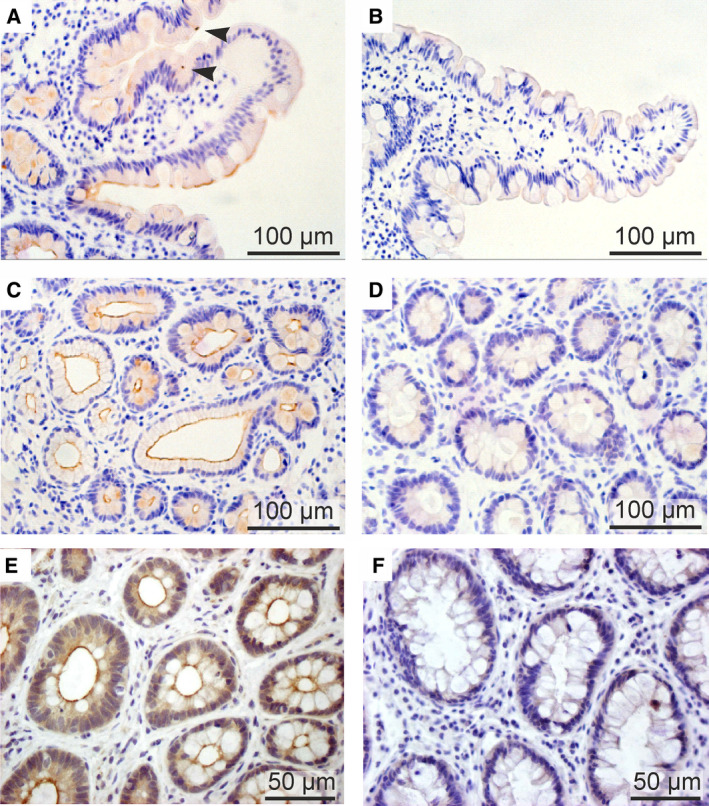
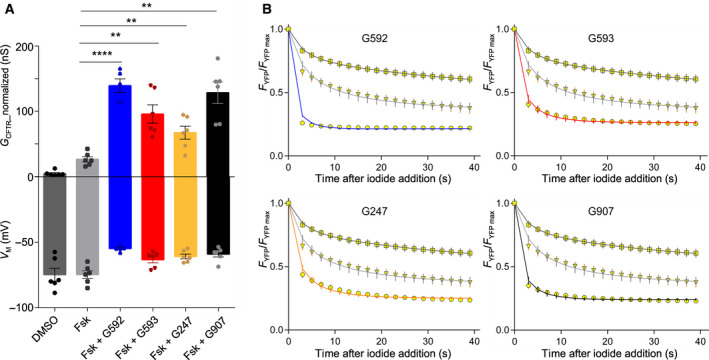
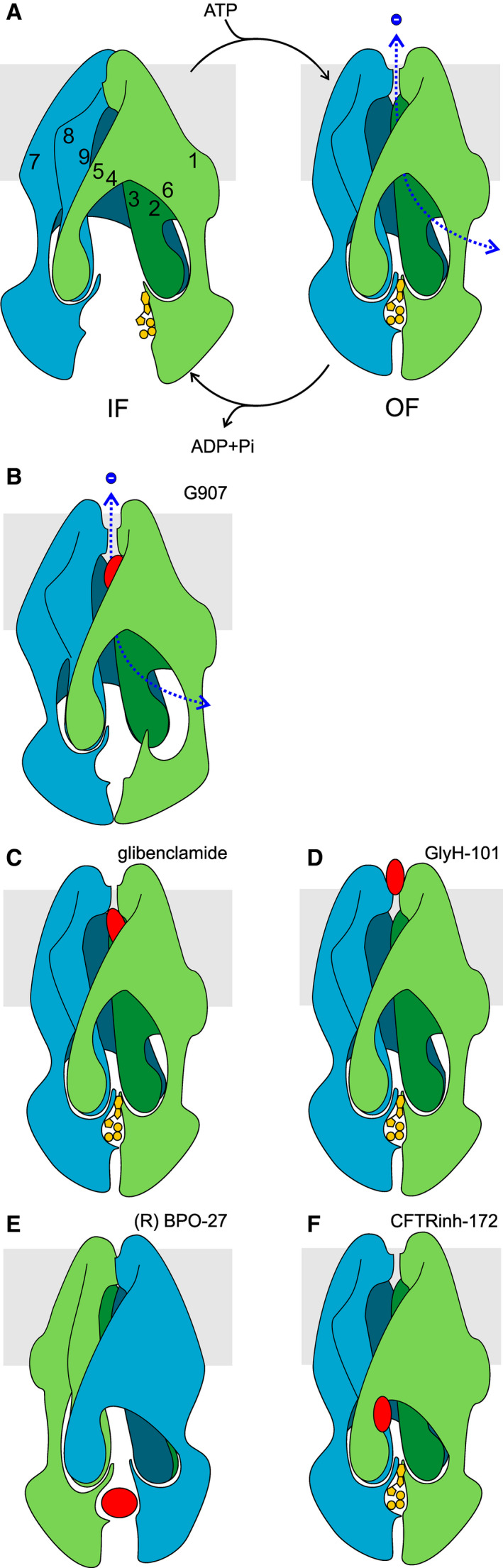
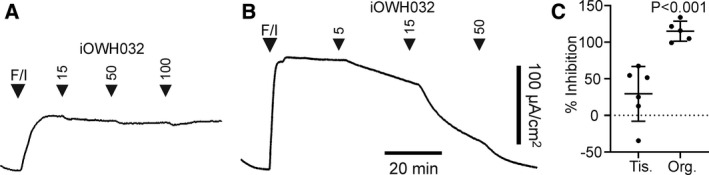
Cystic fibrosis transmembrane conductance regulator (CFTR) is an unusual ABC transporter. It acts as an anion-selective channel that drives osmotic fluid transport across many epithelia. In the gut, CFTR is crucial for maintaining fluid and acid-base homeostasis, and its activity is tightly controlled by multiple neuro-endocrine factors. However, microbial toxins can disrupt this intricate control mechanism and trigger protracted activation of CFTR. This results in the massive faecal water loss, metabolic acidosis and dehydration that characterize secretory diarrhoeas, a major cause of malnutrition and death of children under 5 years of age. Compounds that inhibit CFTR could improve emergency treatment of diarrhoeal disease. Drawing on recent structural and functional insight, we discuss how existing CFTR inhibitors function at the molecular and cellular level. We compare their mechanisms of action to those of inhibitors of related ABC transporters, revealing some unexpected features of drug action on CFTR. Although challenges remain, especially relating to the practical effectiveness of currently available CFTR inhibitors, we discuss how recent technological advances might help develop therapies to better address this important global health need.
Keywords: CFTR pharmacology; G907 compound; cholera; cyclic AMP; cyclic GMP; enterocyte; glibenclamide; ion-channel gating; secretory diarrhea; zosuquidar.
© 2020 The Authors. FEBS Letters published by John Wiley & Sons Ltd on behalf of Federation of European Biochemical Societies.
Figures
References
-
- Csanády L, Vergani P and Gadsby DC (2019) Structure, gating and regulation of the CFTR anion channel. Physiol Rev 99, 707–738. - PubMed
-
- De Boeck K (2020) Cystic fibrosis in the year 2020: a disease with a new face. Acta Paediatr 109, 893–899. - PubMed
-
- Thiagarajah JR, Broadbent T, Hsieh E and Verkman AS (2004) Prevention of toxin‐induced intestinal ion and fluid secretion by a small‐molecule CFTR inhibitor. Gastroenterology 126, 511–519. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
