

Virus-Host Protein-Protein Interactions between Human Papillomavirus 16 E6 A1 and D2/D3 Sub-Lineages: Variances and Similarities
- PMID: 33121134
- PMCID: PMC7663357
- DOI: 10.3390/ijms21217980
Virus-Host Protein-Protein Interactions between Human Papillomavirus 16 E6 A1 and D2/D3 Sub-Lineages: Variances and Similarities
Abstract
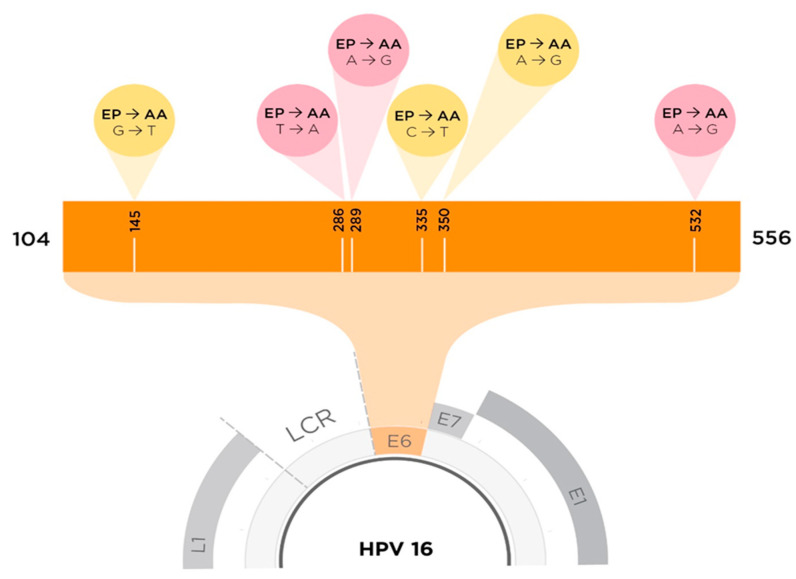
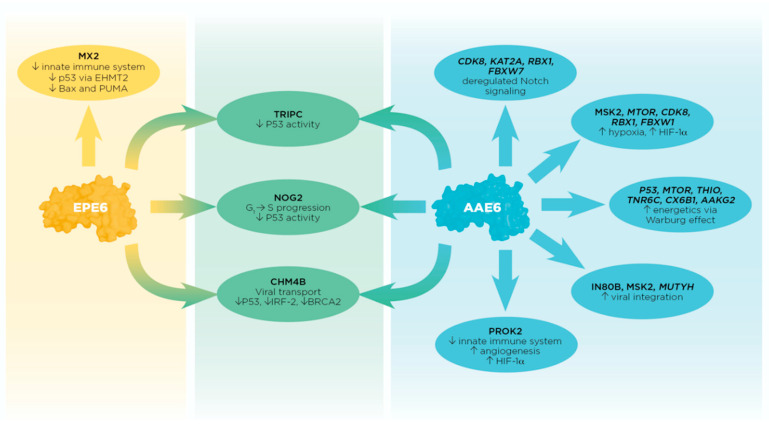
High-risk strains of human papillomavirus are causative agents for cervical and other mucosal cancers, with type 16 being the most frequent. Compared to the European Prototype (EP; A1), the Asian-American (AA; D2/D3) sub-lineage seems to have increased abilities to promote carcinogenesis. Here, we studied protein-protein interactions (PPIs) between host proteins and sub-lineages of the key transforming E6 protein. We transduced human keratinocyte with EP or AA E6 genes and co-immunoprecipitated E6 proteins along with interacting cellular proteins to detect virus-host binding partners. AAE6 and EPE6 may have unique PPIs with host cellular proteins, conferring gain or loss of function and resulting in varied abilities to promote carcinogenesis. Using liquid chromatography-mass spectrometry and stringent interactor selection criteria based on the number of peptides, we identified 25 candidates: 6 unique to AAE6 and EPE6, along with 13 E6 targets common to both. A novel approach based on pathway selection discovered 171 target proteins: 90 unique AAE6 and 61 unique EPE6 along with 20 common E6 targets. Interpretations were made using databases, such as UniProt, BioGRID, and Reactome. Detected E6 targets were differentially implicated in important hallmarks of cancer: deregulating Notch signaling, energetics and hypoxia, DNA replication and repair, and immune response.
Keywords: E6 oncoprotein; carcinogenesis; co-immunoprecipitation; human papillomavirus; interactome study; mass spectrometry; metabolic pathways; proteomics; sub-lineages.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures

Similar articles
-
Two common variants of human papillomavirus type 16 E6 differentially deregulate sugar metabolism and hypoxia signalling in permissive human keratinocytes.J Gen Virol. 2017 Sep;98(9):2310-2319. doi: 10.1099/jgv.0.000905. Epub 2017 Aug 31. J Gen Virol. 2017. PMID: 28857035
-
Tumourigenesis driven by the human papillomavirus type 16 Asian-American e6 variant in a three-dimensional keratinocyte model.PLoS One. 2014 Jul 1;9(7):e101540. doi: 10.1371/journal.pone.0101540. eCollection 2014. PLoS One. 2014. PMID: 24983759 Free PMC article.
-
Functional variants of human papillomavirus type 16 demonstrate host genome integration and transcriptional alterations corresponding to their unique cancer epidemiology.BMC Genomics. 2016 Nov 2;17(1):851. doi: 10.1186/s12864-016-3203-3. BMC Genomics. 2016. PMID: 27806689 Free PMC article.
-
The human papillomavirus 16 European-T350G E6 variant can immortalize but not transform keratinocytes in the absence of E7.Virology. 2015 Nov;485:274-82. doi: 10.1016/j.virol.2015.07.025. Epub 2015 Aug 27. Virology. 2015. PMID: 26318249
-
The Asian-American E6 variant protein of human papillomavirus 16 alone is sufficient to promote immortalization, transformation, and migration of primary human foreskin keratinocytes.J Virol. 2012 Nov;86(22):12384-96. doi: 10.1128/JVI.01512-12. Epub 2012 Sep 5. J Virol. 2012. PMID: 22951839 Free PMC article.
Cited by
-
Metabolic Reprogramming in Cancer: Role of HPV 16 Variants.Pathogens. 2021 Mar 16;10(3):347. doi: 10.3390/pathogens10030347. Pathogens. 2021. PMID: 33809480 Free PMC article. Review.
-
Host-Pathogen Interaction 3.0.Int J Mol Sci. 2022 Oct 24;23(21):12811. doi: 10.3390/ijms232112811. Int J Mol Sci. 2022. PMID: 36361600 Free PMC article.
References
-
- Walboomers J.M.M., Jacobs M.V., Manos M.M., Bosch F.X., Kummer J.A., Shah K.V., Snijders P.J.F., Peto J., Meijer C.J.L.M., Muñoz N. Human papillomavirus is a necessary cause of invasive cervical cancer worldwide. J. Pathol. 1999;189:12–19. doi: 10.1002/(SICI)1096-9896(199909)189:1<12::AID-PATH431>3.0.CO;2-F. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
