

[4Fe-4S] cluster trafficking mediated by Arabidopsis mitochondrial ISCA and NFU proteins
- PMID: 33122194
- PMCID: PMC7939391
- DOI: 10.1074/jbc.RA120.015726
[4Fe-4S] cluster trafficking mediated by Arabidopsis mitochondrial ISCA and NFU proteins
Abstract
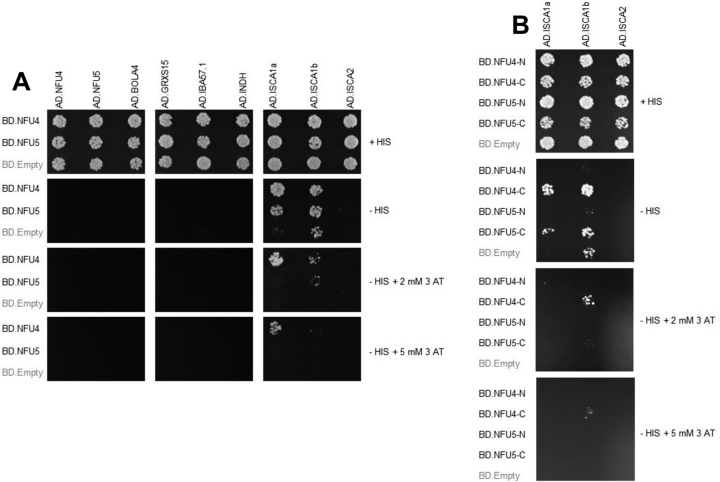
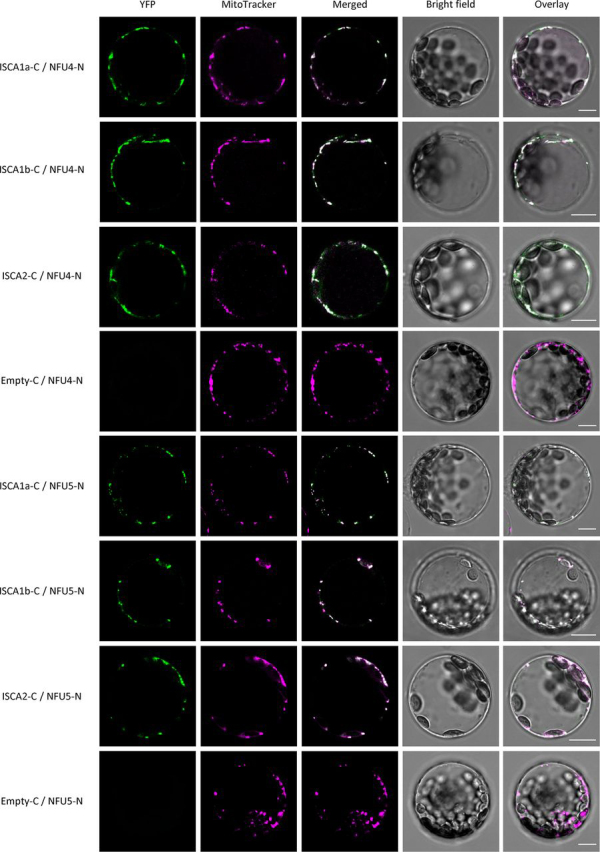
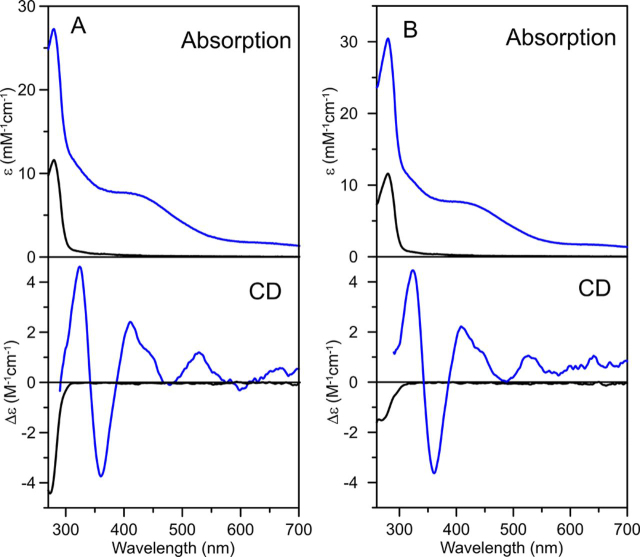
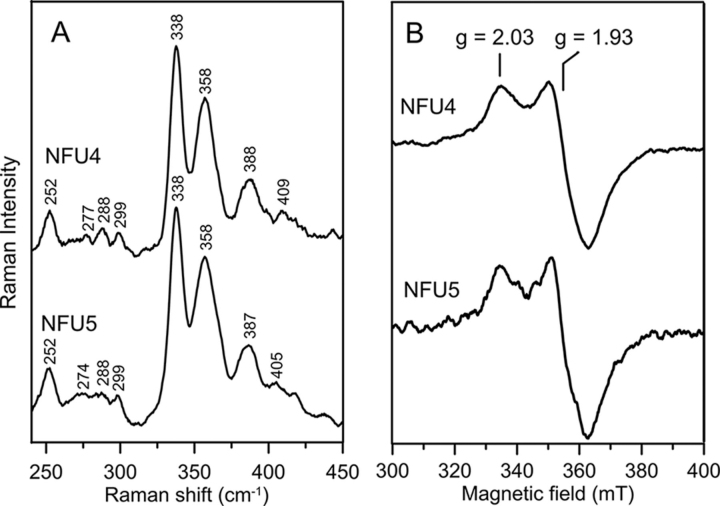
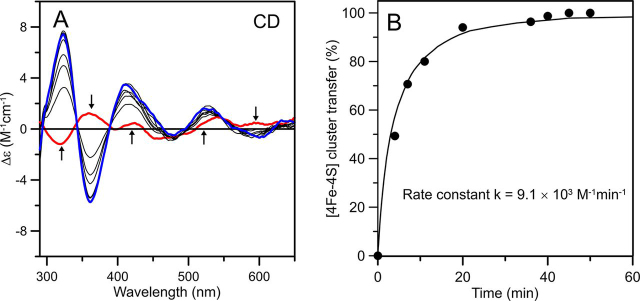
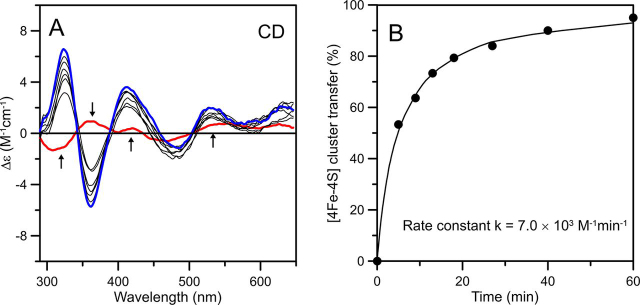
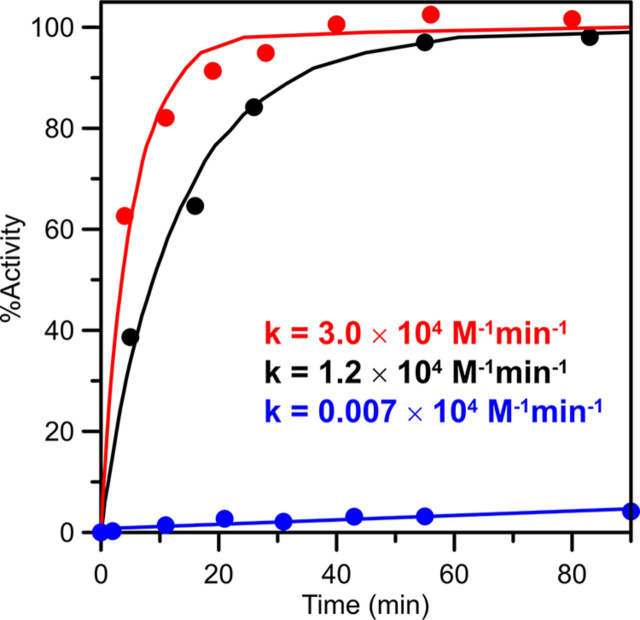
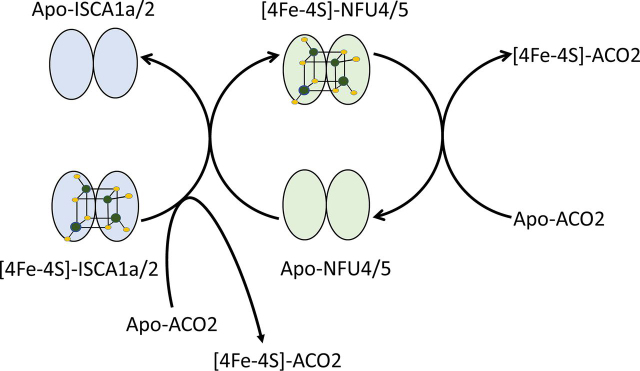
Numerous iron-sulfur (Fe-S) proteins with diverse functions are present in the matrix and respiratory chain complexes of mitochondria. Although [4Fe-4S] clusters are the most common type of Fe-S cluster in mitochondria, the molecular mechanism of [4Fe-4S] cluster assembly and insertion into target proteins by the mitochondrial iron-sulfur cluster (ISC) maturation system is not well-understood. Here we report a detailed characterization of two late-acting Fe-S cluster-carrier proteins from Arabidopsis thaliana, NFU4 and NFU5. Yeast two-hybrid and bimolecular fluorescence complementation studies demonstrated interaction of both the NFU4 and NFU5 proteins with the ISCA class of Fe-S carrier proteins. Recombinant NFU4 and NFU5 were purified as apo-proteins after expression in Escherichia coliIn vitro Fe-S cluster reconstitution led to the insertion of one [4Fe-4S]2+ cluster per homodimer as determined by UV-visible absorption/CD, resonance Raman and EPR spectroscopy, and analytical studies. Cluster transfer reactions, monitored by UV-visible absorption and CD spectroscopy, showed that a [4Fe-4S]2+ cluster-bound ISCA1a/2 heterodimer is effective in transferring [4Fe-4S]2+ clusters to both NFU4 and NFU5 with negligible back reaction. In addition, [4Fe-4S]2+ cluster-bound ISCA1a/2, NFU4, and NFU5 were all found to be effective [4Fe-4S]2+ cluster donors for maturation of the mitochondrial apo-aconitase 2 as assessed by enzyme activity measurements. The results demonstrate rapid, unidirectional, and quantitative [4Fe-4S]2+ cluster transfer from ISCA1a/2 to NFU4 or NFU5 that further delineates their respective positions in the plant ISC machinery and their contributions to the maturation of client [4Fe-4S] cluster-containing proteins.
Keywords: Arabidopsis thaliana; ISCA proteins; NFU proteins; Raman spectroscopy; circular dichroism; iron-sulfur cluster trafficking; iron-sulfur protein; mitochondria; protein-protein interaction.
© 2020 Azam et al.
Conflict of interest statement
Conflict of interest—The authors declare that they have no conflicts of interest with the content of this article.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
