

Replication stress conferred by POT1 dysfunction promotes telomere relocalization to the nuclear pore
- PMID: 33122293
- PMCID: PMC7706707
- DOI: 10.1101/gad.337287.120
Replication stress conferred by POT1 dysfunction promotes telomere relocalization to the nuclear pore
Abstract
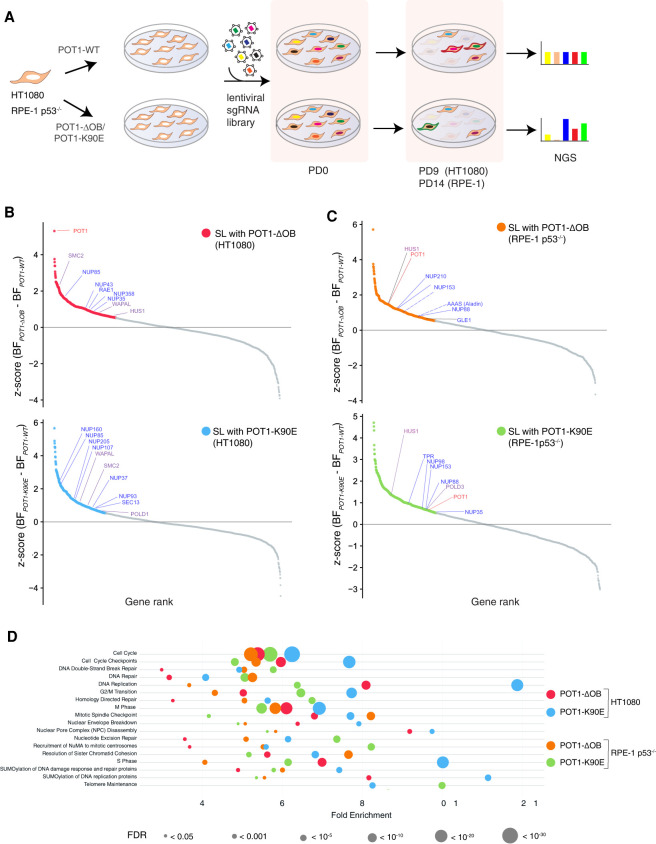
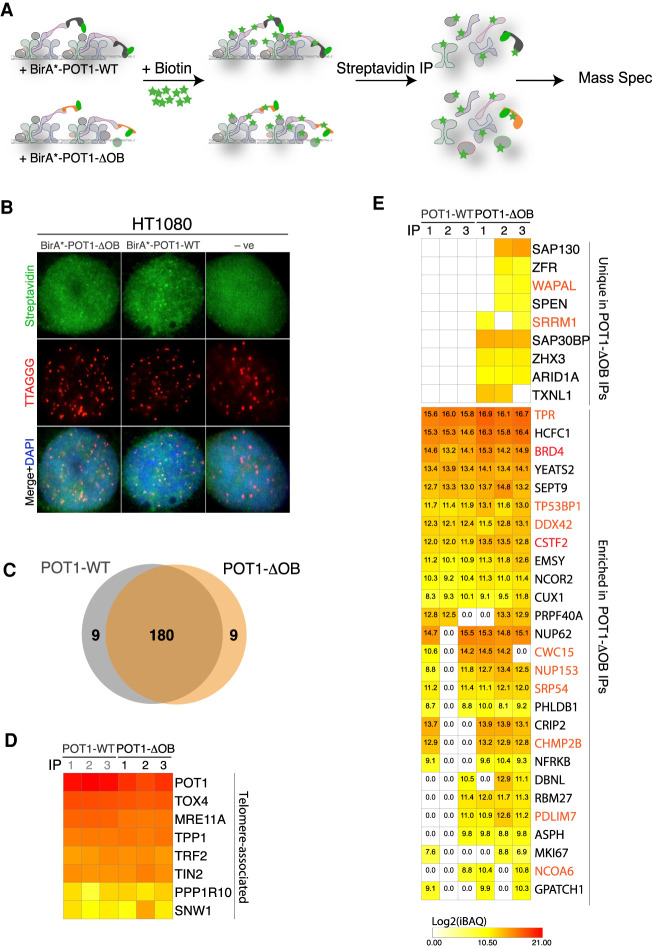
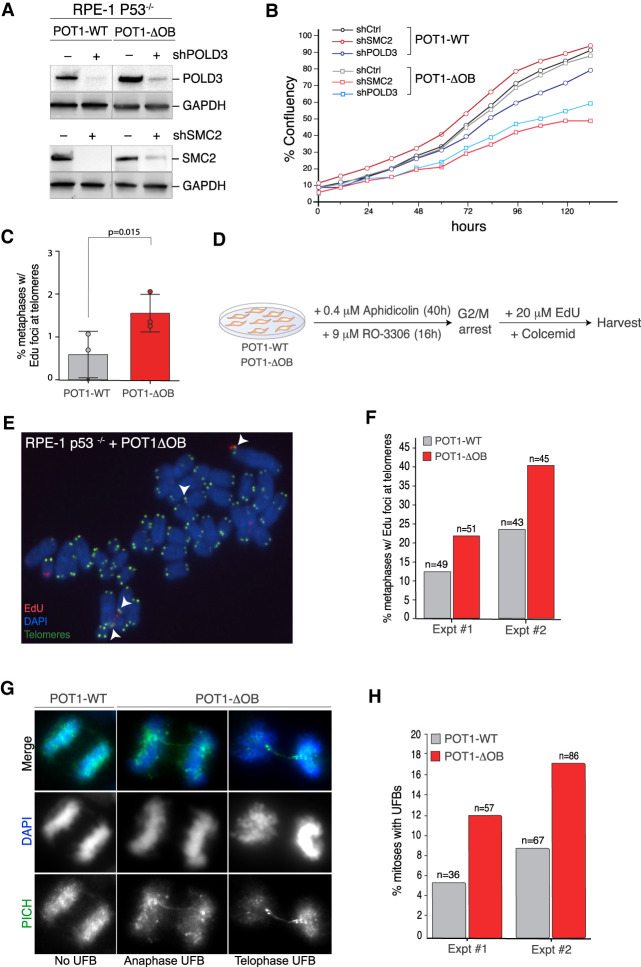
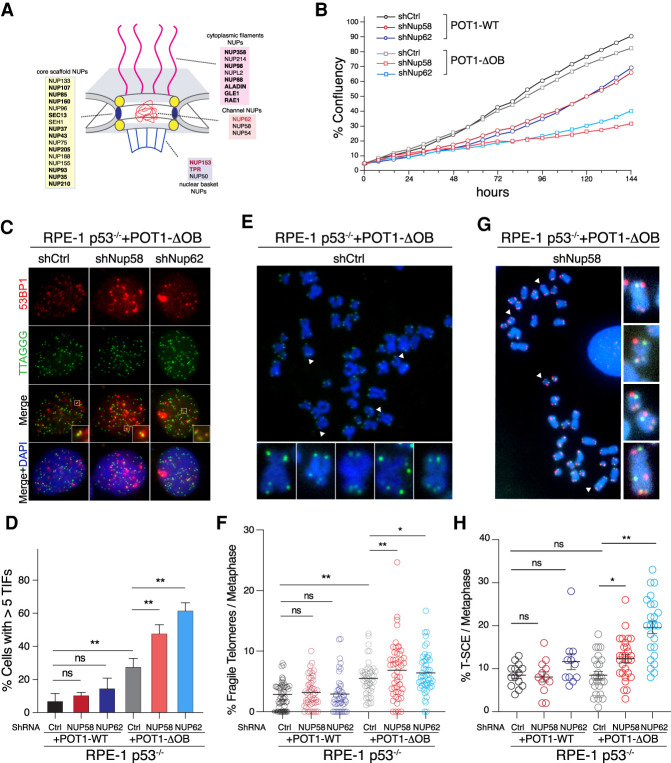
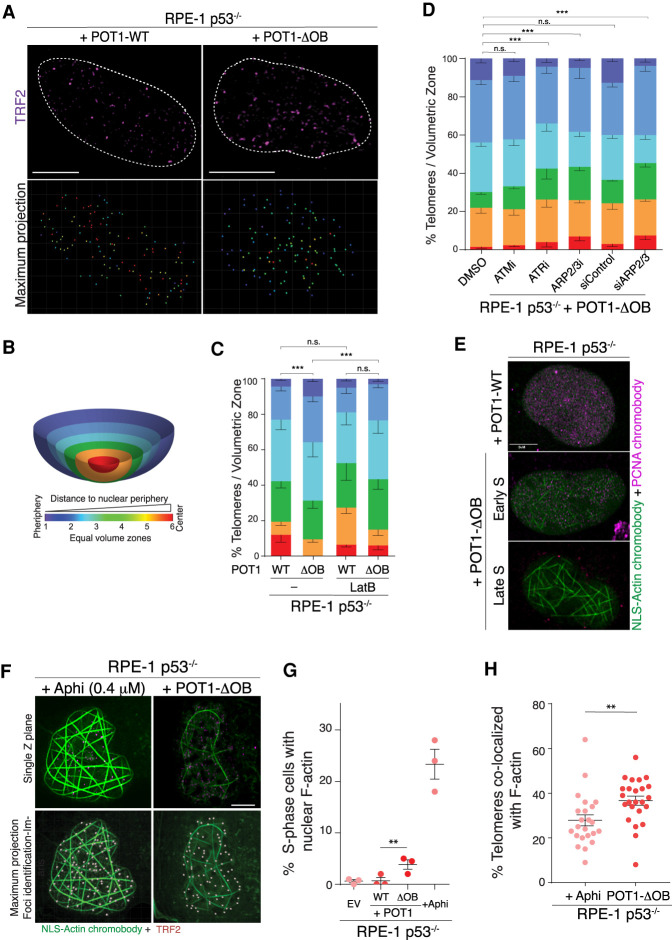
Mutations in the telomere-binding protein POT1 are associated with solid tumors and leukemias. POT1 alterations cause rapid telomere elongation, ATR kinase activation, telomere fragility, and accelerated tumor development. Here, we define the impact of mutant POT1 alleles through complementary genetic and proteomic approaches based on CRISPR interference and biotin-based proximity labeling, respectively. These screens reveal that replication stress is a major vulnerability in cells expressing mutant POT1, which manifests as increased telomere mitotic DNA synthesis at telomeres. Our study also unveils a role for the nuclear pore complex in resolving replication defects at telomeres. Depletion of nuclear pore complex subunits in the context of POT1 dysfunction increases DNA damage signaling, telomere fragility and sister chromatid exchanges. Furthermore, we observed telomere repositioning to the nuclear periphery driven by nuclear F-actin polymerization in cells with POT1 mutations. In conclusion, our study establishes that relocalization of dysfunctional telomeres to the nuclear periphery is critical to preserve telomere repeat integrity.
Keywords: CRISPRi; MiDAS; POT1; nuclear F-actin; nuclear periphery; nuclear pore; replication stress; telomeres.
© 2020 Pinzaru et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous