

Molecular Characterization of a Novel Shell Matrix Protein With PDZ Domain From Mytilus coruscus
- PMID: 33123020
- PMCID: PMC7573561
- DOI: 10.3389/fphys.2020.543758
Molecular Characterization of a Novel Shell Matrix Protein With PDZ Domain From Mytilus coruscus
Abstract
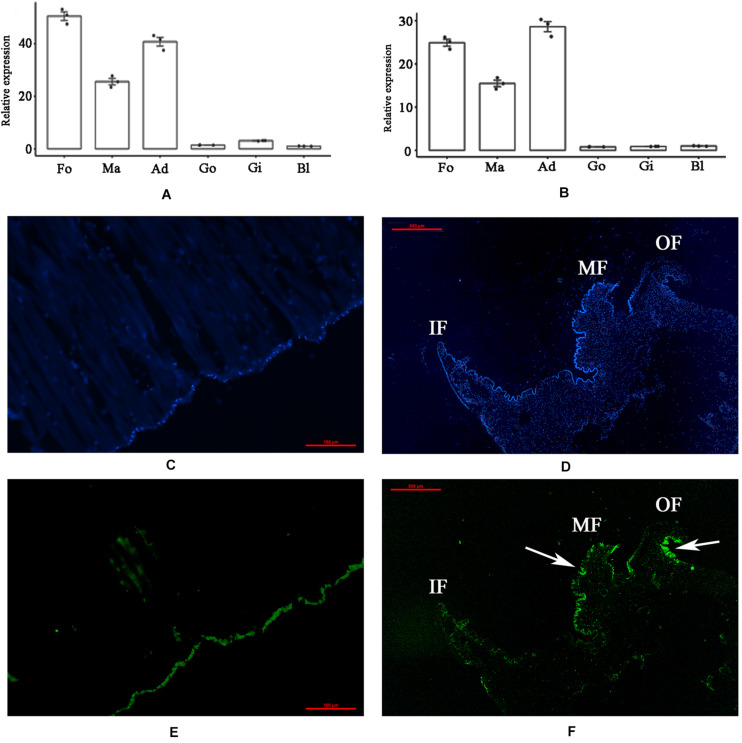
Mollusk shells are products of biomineralization and possess excellent mechanical properties, and shell matrix proteins (SMPs) have important functions in shell formation. A novel SMP with a PDZ domain (PDZ-domain-containing-protein-1, PDCP-1) was identified from the shell matrices of Mytilus coruscus. In this study, the gene expression, function, and location of PDCP-1 were analyzed. PDCP-1 was characterized as an ∼70 kDa protein with a PDZ (postsynaptic density/discs large/zonula occludes) domain and a ZM (ZASP-like motif) domain. The PDCP-1 gene has a high expression level and specific location in the foot, mantle and adductor muscle. Recombinantly expressed PDCP-1 (rPDCP-1) altered the morphology of calcite crystals, the polymorph of calcite crystals, binding with both calcite and aragonite crystals, and inhibition of the crystallization rate of calcite crystals. In addition, anti-rPDCP-1 antibody was prepared, and immunohistochemistry and immunofluorescence analyses revealed the specific location of PDCP-1 in the mantle, the adductor muscle, and the aragonite (nacre and myostracum) layer of the shell, suggesting multiple functions of PDCP-1 in biomineralization, muscle-shell attachment, and muscle attraction. Furthermore, pull-down analysis revealed 19 protein partners of PDCP-1 from the shell matrices, which accordingly provided a possible interaction network of PDCP-1 in the shell. These results expand the understanding of the functions of PDZ-domain-containing proteins (PDCPs) in biomineralization and the supramolecular chemistry that contributes to shell formation.
Keywords: Mytilus coruscus; PDZ-domain-containing protein; biomineralization; recombinant expression; shell matrix proteins.
Copyright © 2020 Sun, Jiang, Yan, Fan, Zhang, Xu and Liao.
Figures







References
-
- Addadi L., Weiner S. (1986). Interactions between acidic macromolecules and structured crystal surfaces. Stereochemistry and biomineralization. Mol. Crys. Liquid Crystals 134 305–322. 10.1080/00268948608079592 - DOI
LinkOut - more resources
Full Text Sources
Research Materials