

Transcriptome and epigenome diversity and plasticity of muscle stem cells following transplantation
- PMID: 33125370
- PMCID: PMC7657492
- DOI: 10.1371/journal.pgen.1009022
Transcriptome and epigenome diversity and plasticity of muscle stem cells following transplantation
Abstract
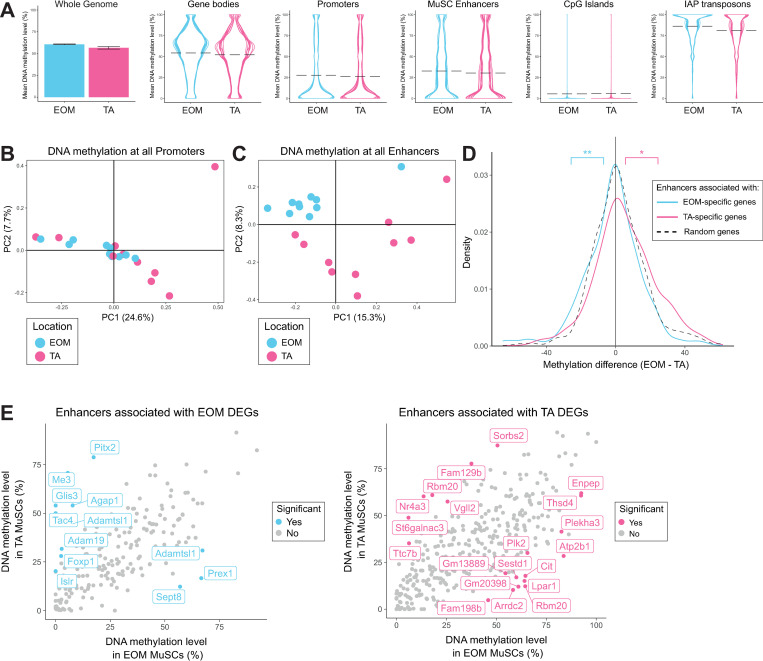
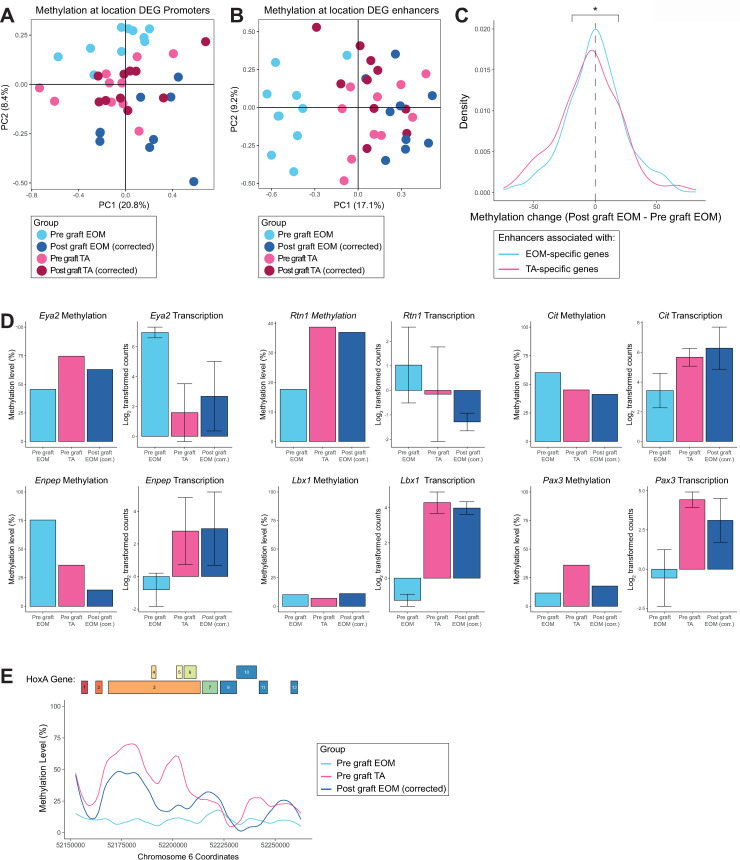
Adult skeletal muscles are maintained during homeostasis and regenerated upon injury by muscle stem cells (MuSCs). A heterogeneity in self-renewal, differentiation and regeneration properties has been reported for MuSCs based on their anatomical location. Although MuSCs derived from extraocular muscles (EOM) have a higher regenerative capacity than those derived from limb muscles, the molecular determinants that govern these differences remain undefined. Here we show that EOM and limb MuSCs have distinct DNA methylation signatures associated with enhancers of location-specific genes, and that the EOM transcriptome is reprogrammed following transplantation into a limb muscle environment. Notably, EOM MuSCs expressed host-site specific positional Hox codes after engraftment and self-renewal within the host muscle. However, about 10% of EOM-specific genes showed engraftment-resistant expression, pointing to cell-intrinsic molecular determinants of the higher engraftment potential of EOM MuSCs. Our results underscore the molecular diversity of distinct MuSC populations and molecularly define their plasticity in response to microenvironmental cues. These findings provide insights into strategies designed to improve the functional capacity of MuSCs in the context of regenerative medicine.
Conflict of interest statement
I have read the journal's policy and the authors of this manuscript have the following competing interests: W.R. is a consultant and shareholder of Cambridge Epigenetix. T.S. is CEO and shareholder of Chronomics. All other authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
