

Role of Polycomb Complexes in Normal and Malignant Plasma Cells
- PMID: 33126754
- PMCID: PMC7662980
- DOI: 10.3390/ijms21218047
Role of Polycomb Complexes in Normal and Malignant Plasma Cells
Abstract
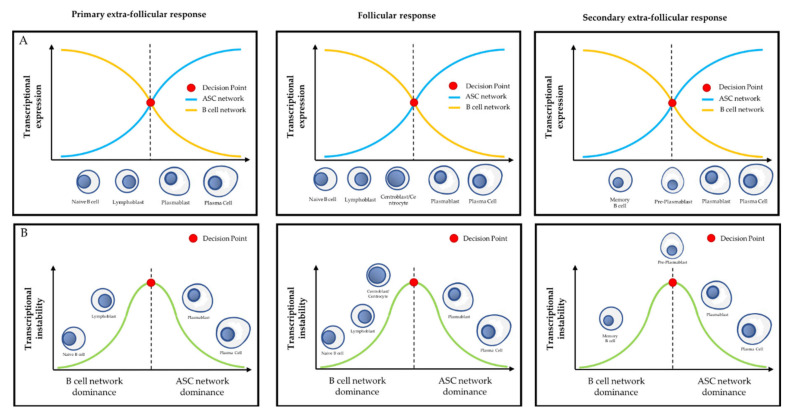
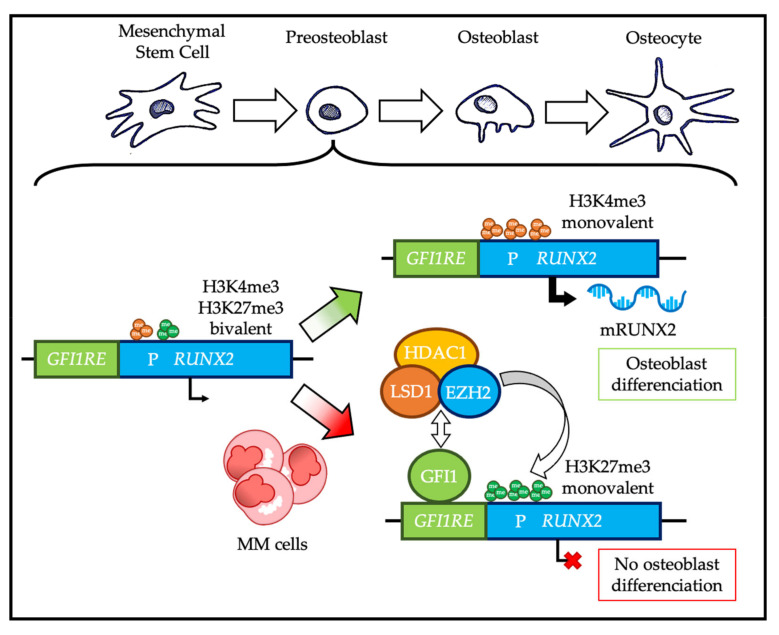
Plasma cells (PC) are the main effectors of adaptive immunity, responsible for producing antibodies to defend the body against pathogens. They are the result of a complex highly regulated cell differentiation process, taking place in several anatomical locations and involving unique genetic events. Pathologically, PC can undergo tumorigenesis and cause a group of diseases known as plasma cell dyscrasias, including multiple myeloma (MM). MM is a severe disease with poor prognosis that is characterized by the accumulation of malignant PC within the bone marrow, as well as high clinical and molecular heterogeneity. MM patients frequently develop resistance to treatment, leading to relapse. Polycomb group (PcG) proteins are epigenetic regulators involved in cell fate and carcinogenesis. The emerging roles of PcG in PC differentiation and myelomagenesis position them as potential therapeutic targets in MM. Here, we focus on the roles of PcG proteins in normal and malignant plasma cells, as well as their therapeutic implications.
Keywords: epigenetics; multiple myeloma; plasma cell differentiation; polycomb.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
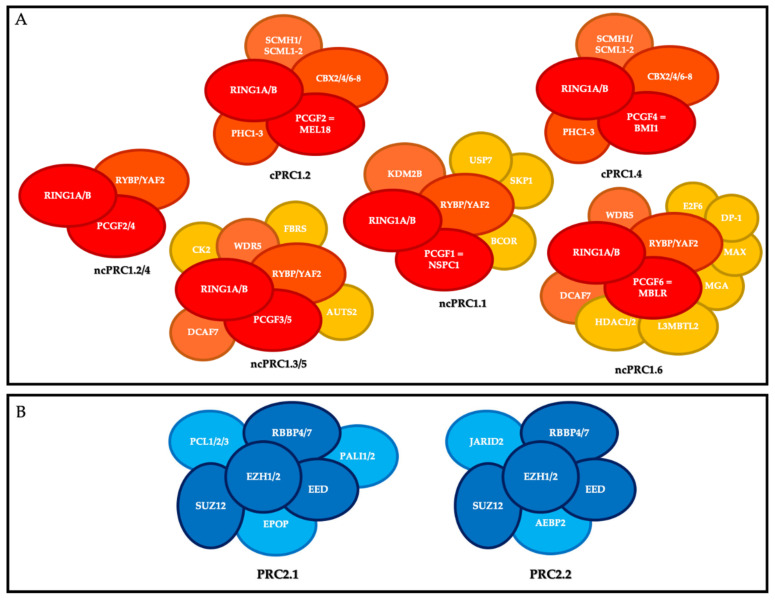
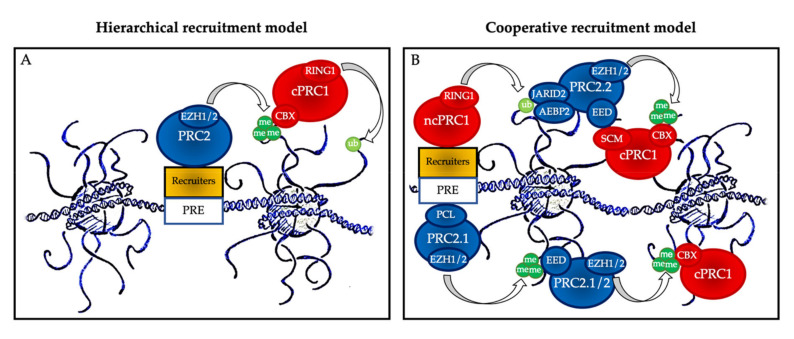
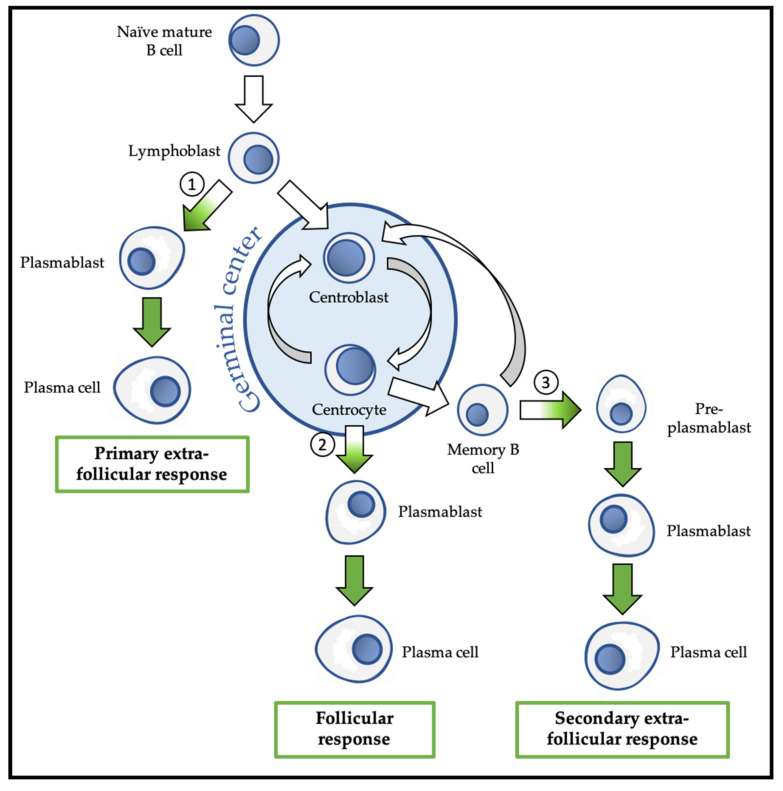
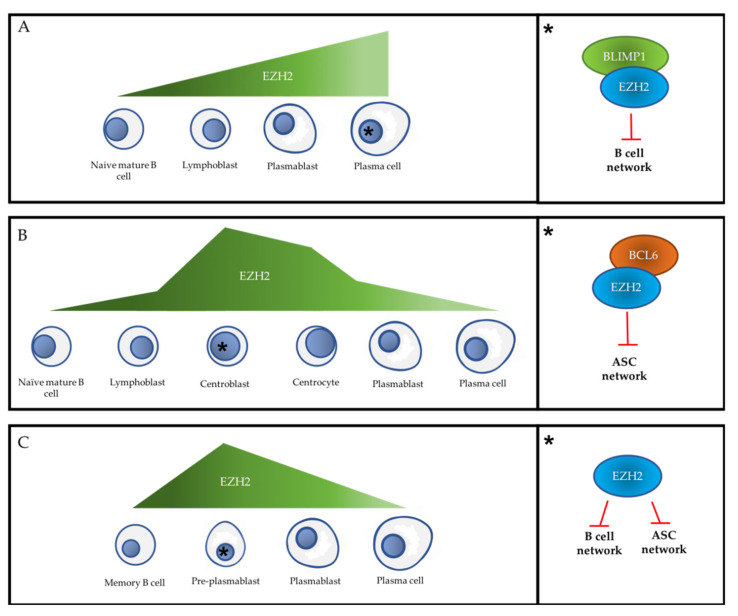
References
-
- Maximow A.A. Experimentelle Untersuchungen über Entzündliche Neubildung von Bindegewebe. Fischer; Leipzig, Germany: 1902. Chapter III-C.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
