

A role of oligodendrocytes in information processing
- PMID: 33127910
- PMCID: PMC7599337
- DOI: 10.1038/s41467-020-19152-7
A role of oligodendrocytes in information processing
Abstract
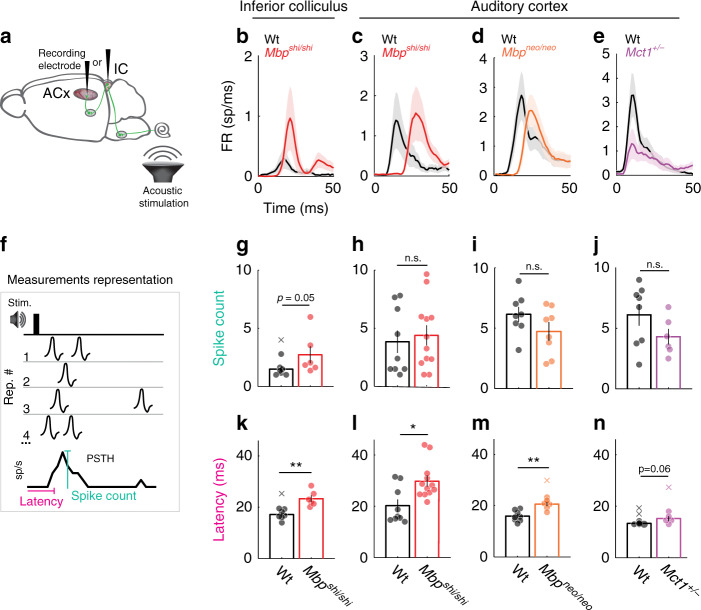
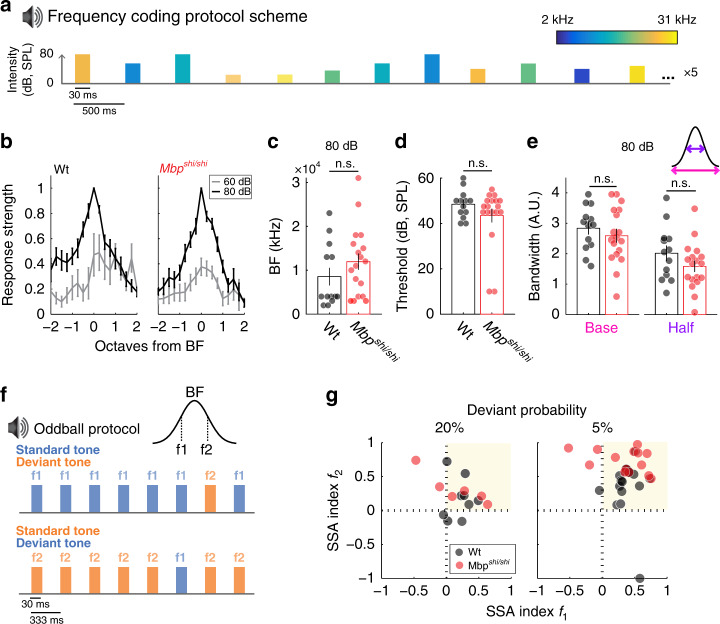
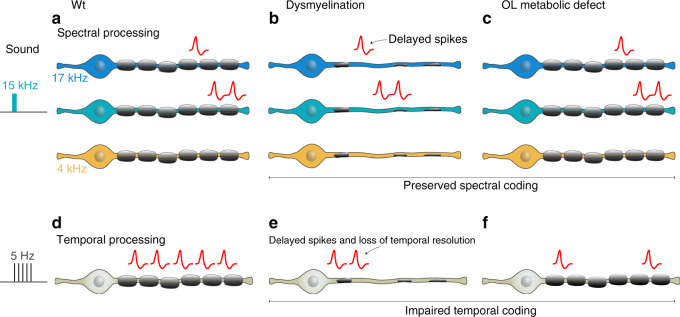
Myelinating oligodendrocytes enable fast propagation of action potentials along the ensheathed axons. In addition, oligodendrocytes play diverse non-canonical roles including axonal metabolic support and activity-dependent myelination. An open question remains whether myelination also contributes to information processing in addition to speeding up conduction velocity. Here, we analyze the role of myelin in auditory information processing using paradigms that are also good predictors of speech understanding in humans. We compare mice with different degrees of dysmyelination using acute multiunit recordings in the auditory cortex, in combination with behavioral readouts. We find complex alterations of neuronal responses that reflect fatigue and temporal acuity deficits. We observe partially discriminable but similar deficits in well myelinated mice in which glial cells cannot fully support axons metabolically. We suggest a model in which myelination contributes to sustained stimulus perception in temporally complex paradigms, with a role of metabolically active oligodendrocytes in cortical information processing.
Conflict of interest statement
The authors declare no competing interests.
Figures
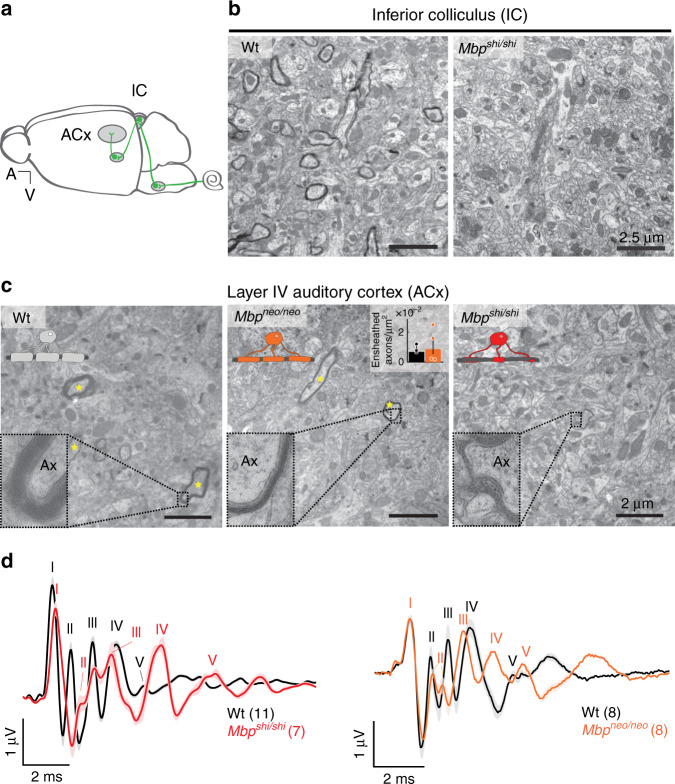
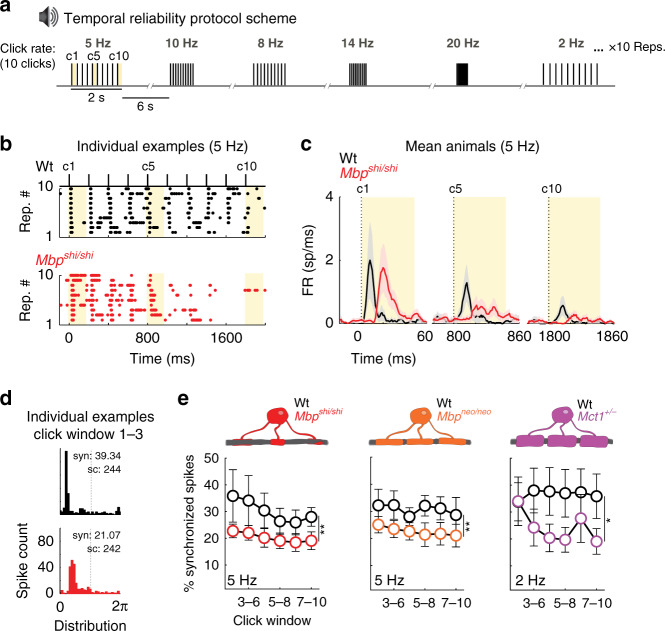
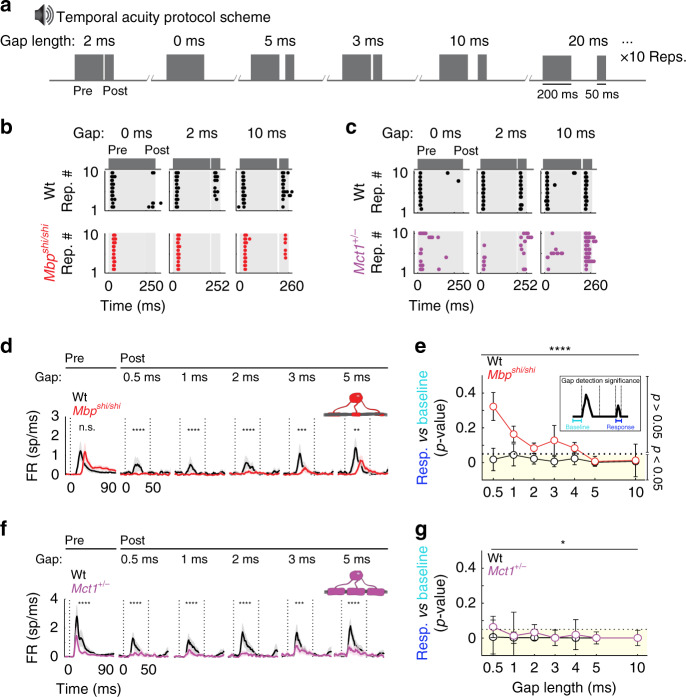
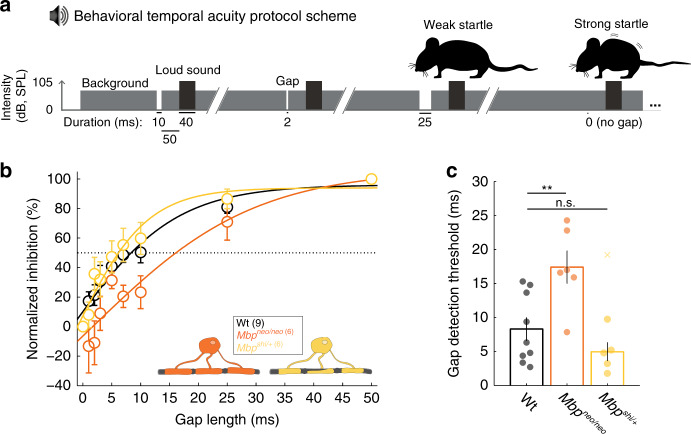
Similar articles
-
Axoglial interactions in myelin plasticity: Evaluating the relationship between neuronal activity and oligodendrocyte dynamics.Glia. 2019 Nov;67(11):2038-2049. doi: 10.1002/glia.23629. Epub 2019 Apr 30. Glia. 2019. PMID: 31038804 Review.
-
Oligodendrocytes: facilitating axonal conduction by more than myelination.Neuroscientist. 2010 Feb;16(1):11-8. doi: 10.1177/1073858409334425. Epub 2009 May 8. Neuroscientist. 2010. PMID: 19429890 Review.
-
Oligodendrocytes: Myelination and Axonal Support.Cold Spring Harb Perspect Biol. 2015 Jun 22;8(1):a020479. doi: 10.1101/cshperspect.a020479. Cold Spring Harb Perspect Biol. 2015. PMID: 26101081 Free PMC article. Review.
-
Individual Oligodendrocytes Show Bias for Inhibitory Axons in the Neocortex.Cell Rep. 2019 Jun 4;27(10):2799-2808.e3. doi: 10.1016/j.celrep.2019.05.018. Cell Rep. 2019. PMID: 31167127 Free PMC article.
-
The acquisition of myelin: a success story.Novartis Found Symp. 2006;276:15-21; discussion 21-5, 54-7, 275-81. doi: 10.1002/9780470032244.ch3. Novartis Found Symp. 2006. PMID: 16805421 Review.
Cited by
-
Motor learning drives dynamic patterns of intermittent myelination on learning-activated axons.Nat Neurosci. 2022 Oct;25(10):1300-1313. doi: 10.1038/s41593-022-01169-4. Epub 2022 Sep 30. Nat Neurosci. 2022. PMID: 36180791 Free PMC article.
-
ECM-derived biomaterials for regulating tissue multicellularity and maturation.iScience. 2024 Feb 6;27(3):109141. doi: 10.1016/j.isci.2024.109141. eCollection 2024 Mar 15. iScience. 2024. PMID: 38405613 Free PMC article. Review.
-
Deletion of the Sodium-Dependent Glutamate Transporter GLT-1 in Maturing Oligodendrocytes Attenuates Myelination of Callosal Axons During a Postnatal Phase of Central Nervous System Development.Front Cell Neurosci. 2022 Jun 3;16:905299. doi: 10.3389/fncel.2022.905299. eCollection 2022. Front Cell Neurosci. 2022. PMID: 35722615 Free PMC article.
-
Dynamics of mature myelin.Nat Neurosci. 2024 Aug;27(8):1449-1461. doi: 10.1038/s41593-024-01642-2. Epub 2024 May 21. Nat Neurosci. 2024. PMID: 38773349 Free PMC article. Review.
-
Functional myelin in cognition and neurodevelopmental disorders.Cell Mol Life Sci. 2024 Apr 13;81(1):181. doi: 10.1007/s00018-024-05222-2. Cell Mol Life Sci. 2024. PMID: 38615095 Free PMC article. Review.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
