

Lipid headgroups alter huntingtin aggregation on membranes
- PMID: 33130095
- PMCID: PMC7742038
- DOI: 10.1016/j.bbamem.2020.183497
Lipid headgroups alter huntingtin aggregation on membranes
Abstract
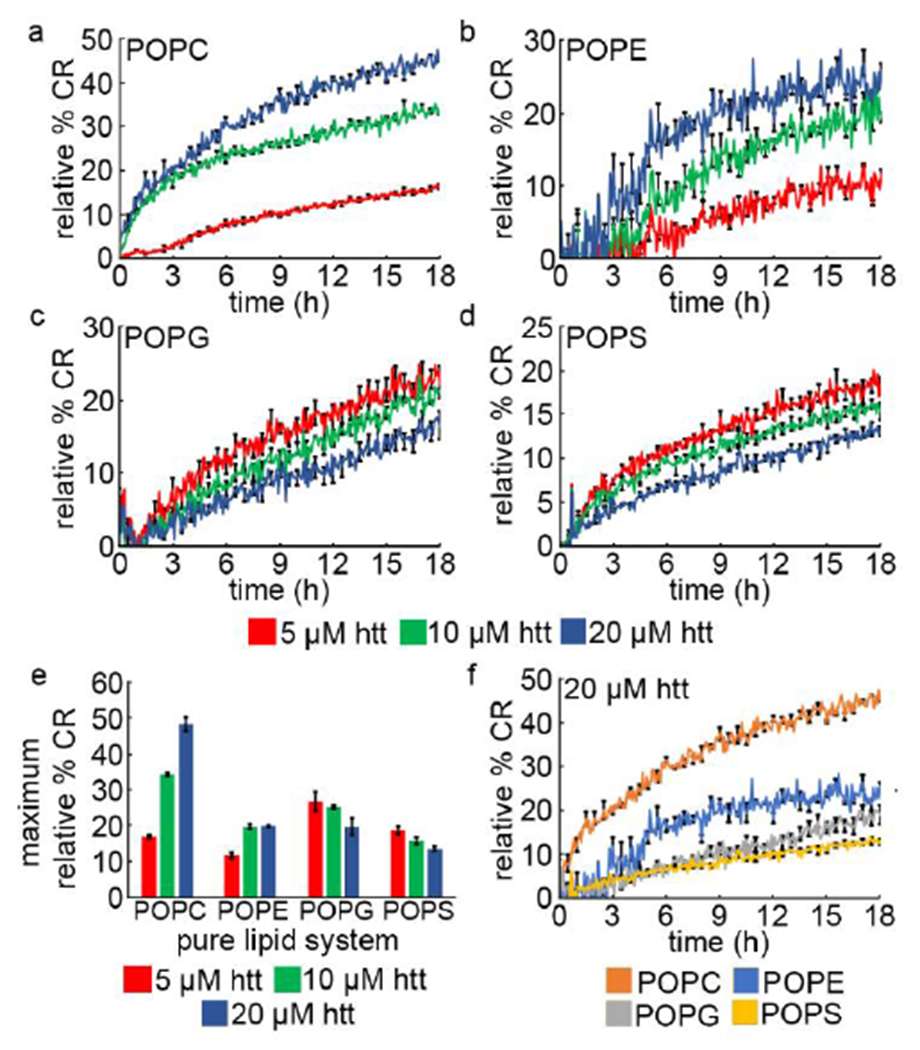
Huntington's Disease is a fatal neurodegenerative disorder caused by expansion of a glutamine repeat region (polyQ) beyond a critical threshold within exon1 of the huntingtin protein (htt). As a consequence of polyQ expansion, htt associates into a variety of aggregate species that are thought to underlie cellular toxicity. Within cells, htt associates with numerous membranous organelles and surfaces that exert influence on the aggregation process. In particular, the first 17 amino acids at the N-terminus of htt (Nt17) serve as a lipid-binding domain that is intrinsically disordered in bulk solution but adopts an amphipathic α-helical structure upon binding membranes. Beyond this, Nt17 is implicated in initiating htt fibrillization. As the interaction between Nt17 and lipid membranes is likely influenced by lipid properties, the impact of lipid headgroups on htt-exon1 aggregation, membrane activity, and the ability to form protein:lipid complexes was determined. Htt-exon1 with a disease-length polyQ domain (46Q) was exposed to lipid vesicles comprised of lipids with either zwitterionic (POPC and POPE) or anionic (POPG and POPS) headgroups. With zwitterionic head groups, large lipid to peptide ratios were required to have a statistically significant impact on htt aggregation. Anionic lipids enhanced htt fibrillization, even at low lipid:protein ratios, and this was accompanied by changes in aggregate morphology. Despite the larger impact of anionic lipids, htt-exon1(46Q) was more membrane active with zwitterionic lipid systems. The ability of Nt17 to form complexes with lipids was also mediated by lipid headgroups as zwitterionic ionic lipids more readily associated with multimeric forms of Nt17 in comparison with anionic lipids. Collectively, these results highlight the complexity of htt/membrane interactions and the resulting impact on the aggregation process.
Keywords: Amyloid; Atomic force microscopy; Electrospray ionization-mass spectrometry; Huntington's disease; Lipid membranes; Polyglutamine.
Copyright © 2020. Published by Elsevier B.V.
Conflict of interest statement
Declaration of competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures






Similar articles
-
SUMOylation Prevents Huntingtin Fibrillization and Localization onto Lipid Membranes.ACS Chem Neurosci. 2020 Feb 5;11(3):328-343. doi: 10.1021/acschemneuro.9b00509. Epub 2020 Jan 9. ACS Chem Neurosci. 2020. PMID: 31880908
-
The interaction of polyglutamine peptides with lipid membranes is regulated by flanking sequences associated with huntingtin.J Biol Chem. 2013 May 24;288(21):14993-5005. doi: 10.1074/jbc.M112.446237. Epub 2013 Apr 9. J Biol Chem. 2013. PMID: 23572526 Free PMC article.
-
Huntingtin disrupts lipid bilayers in a polyQ-length dependent manner.Biochim Biophys Acta. 2013 Aug;1828(8):1953-61. doi: 10.1016/j.bbamem.2013.04.025. Epub 2013 May 2. Biochim Biophys Acta. 2013. PMID: 23643759
-
The emerging role of the first 17 amino acids of huntingtin in Huntington's disease.Biomol Concepts. 2015 Mar;6(1):33-46. doi: 10.1515/bmc-2015-0001. Biomol Concepts. 2015. PMID: 25741791 Free PMC article. Review.
-
Huntingtin interactions with membrane phospholipids: strategic targets for therapeutic intervention?J Huntingtons Dis. 2013;2(3):239-50. doi: 10.3233/JHD-130068. J Huntingtons Dis. 2013. PMID: 25062673 Review.
Cited by
-
The long and the short of Huntington's disease: how the sphingolipid profile is shifted in the caudate of advanced clinical cases.Brain Commun. 2021 Dec 23;4(1):fcab303. doi: 10.1093/braincomms/fcab303. eCollection 2022. Brain Commun. 2021. PMID: 35169703 Free PMC article.
-
The polyglutamine domain is the primary driver of seeding in huntingtin aggregation.PLoS One. 2024 Mar 14;19(3):e0298323. doi: 10.1371/journal.pone.0298323. eCollection 2024. PLoS One. 2024. PMID: 38483973 Free PMC article.
-
Mitochondrial membranes modify mutant huntingtin aggregation.Biochim Biophys Acta Biomembr. 2021 Oct 1;1863(10):183663. doi: 10.1016/j.bbamem.2021.183663. Epub 2021 Jun 2. Biochim Biophys Acta Biomembr. 2021. PMID: 34089719 Free PMC article.
-
The N17 domain of huntingtin as a multifaceted player in Huntington's disease.Front Mol Biosci. 2025 Jan 7;11:1527313. doi: 10.3389/fmolb.2024.1527313. eCollection 2024. Front Mol Biosci. 2025. PMID: 39845903 Free PMC article. Review.
-
Lamellarity-Driven Differences in Surface Structural Features of DPPS Lipids: Spectroscopic, Calorimetric and Computational Study.Membranes (Basel). 2023 Jan 9;13(1):83. doi: 10.3390/membranes13010083. Membranes (Basel). 2023. PMID: 36676890 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
