

Extracellular Histones Inhibit Fibrinolysis through Noncovalent and Covalent Interactions with Fibrin
- PMID: 33131044
- PMCID: PMC7982298
- DOI: 10.1055/s-0040-1718760
Extracellular Histones Inhibit Fibrinolysis through Noncovalent and Covalent Interactions with Fibrin
Abstract
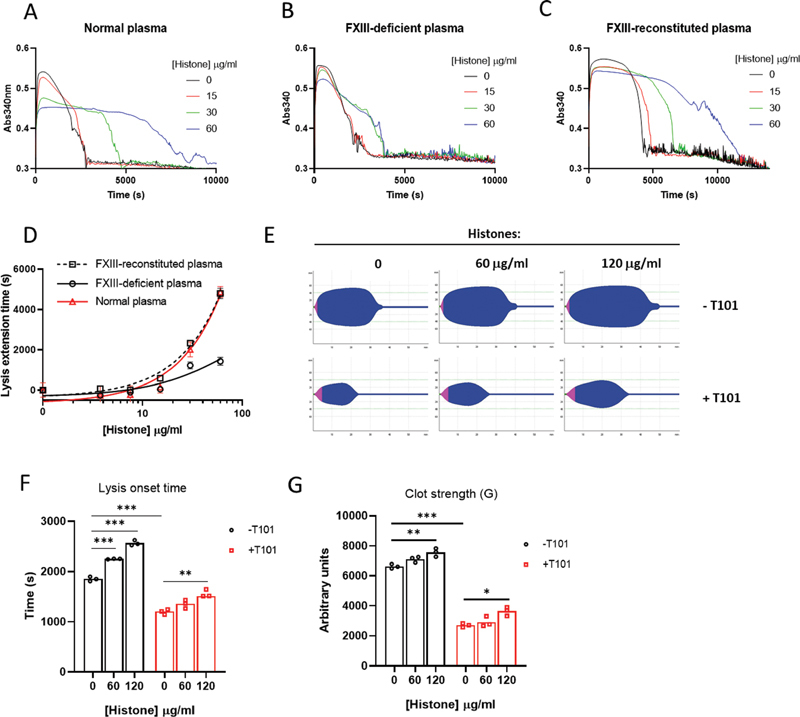
Histones released into circulation as neutrophil extracellular traps are causally implicated in the pathogenesis of arterial, venous, and microvascular thrombosis by promoting coagulation and enhancing clot stability. Histones induce structural changes in fibrin rendering it stronger and resistant to fibrinolysis. The current study extends these observations by defining the antifibrinolytic mechanisms of histones in purified, plasma, and whole blood systems. Although histones stimulated plasminogen activation in solution, they inhibited plasmin as competitive substrates. Protection of fibrin from plasmin digestion is enhanced by covalent incorporation of histones into fibrin, catalyzed by activated transglutaminase, coagulation factor FXIII (FXIIIa). All histone subtypes (H1, H2A, H2B, H3, and H4) were crosslinked to fibrin. A distinct, noncovalent mechanism explains histone-accelerated lateral aggregation of fibrin protofibrils, resulting in thicker fibers with higher mass-to-length ratios and in turn hampered fibrinolysis. However, histones were less effective at delaying fibrinolysis in the absence of FXIIIa activity. Therapeutic doses of low-molecular-weight heparin (LMWH) prevented covalent but not noncovalent histone-fibrin interactions and neutralized the effects of histones on fibrinolysis. This suggests an additional antithrombotic mechanism for LMWH beyond anticoagulation. In conclusion, for the first time we report that histones are crosslinked to fibrin by FXIIIa and promote fibrinolytic resistance which can be overcome by FXIIIa inhibitors and histone-binding heparinoids. These findings provide a rationale for targeting the FXIII-histone-fibrin axis to destabilize fibrin and prevent potentially thrombotic fibrin networks.
Thieme. All rights reserved.
Conflict of interest statement
None declared.
Figures





Similar articles
-
DNA, histones and neutrophil extracellular traps exert anti-fibrinolytic effects in a plasma environment.Thromb Haemost. 2015 Jun;113(6):1289-98. doi: 10.1160/TH14-08-0669. Epub 2015 Mar 19. Thromb Haemost. 2015. PMID: 25789443
-
Kringle-Dependent Inhibition of Plasmin-Mediated Fibrinolysis by Native and Citrullinated Core Histones.Int J Mol Sci. 2025 Jun 17;26(12):5799. doi: 10.3390/ijms26125799. Int J Mol Sci. 2025. PMID: 40565263 Free PMC article.
-
Size- and charge-dependent modulation of the lytic susceptibility and mechanical stability of fibrin-histone clots by heparin and polyphosphate variants.J Thromb Haemost. 2021 May;19(5):1307-1318. doi: 10.1111/jth.15258. Epub 2021 Feb 20. J Thromb Haemost. 2021. PMID: 33609065
-
Factor XIII and venous thromboembolism.Semin Thromb Hemost. 2011 Apr;37(3):305-14. doi: 10.1055/s-0031-1273094. Epub 2011 Mar 31. Semin Thromb Hemost. 2011. PMID: 21455864 Review.
-
The fibrinolytic system in man.Crit Rev Oncol Hematol. 1986;4(3):249-301. doi: 10.1016/s1040-8428(86)80014-2. Crit Rev Oncol Hematol. 1986. PMID: 2420482 Review.
Cited by
-
A Modified in vitro Clot Lysis Assay Predicts Outcomes in Non-traumatic Intracerebral Hemorrhage Stroke Patients-The IRONHEART Study.Front Neurol. 2021 Apr 20;12:613441. doi: 10.3389/fneur.2021.613441. eCollection 2021. Front Neurol. 2021. PMID: 33959087 Free PMC article.
-
Location, location, location: Fibrin, cells, and fibrinolytic factors in thrombi.Front Cardiovasc Med. 2023 Jan 18;9:1070502. doi: 10.3389/fcvm.2022.1070502. eCollection 2022. Front Cardiovasc Med. 2023. PMID: 36741833 Free PMC article. Review.
-
Disseminated intravascular coagulation.J Intensive Care. 2025 Jun 6;13(1):32. doi: 10.1186/s40560-025-00794-y. J Intensive Care. 2025. PMID: 40474215 Free PMC article. Review.
-
The Emerging Role of Neutrophil Extracellular Traps in Arterial, Venous and Cancer-Associated Thrombosis.Front Cardiovasc Med. 2021 Dec 2;8:786387. doi: 10.3389/fcvm.2021.786387. eCollection 2021. Front Cardiovasc Med. 2021. PMID: 34926629 Free PMC article. Review.
-
Blood Clot Dynamics and Fibrinolysis Impairment in Cancer: The Role of Plasma Histones and DNA.Cancers (Basel). 2024 Feb 25;16(5):928. doi: 10.3390/cancers16050928. Cancers (Basel). 2024. PMID: 38473289 Free PMC article.
References
-
- Brinkmann V, Reichard U, Goosmann C.Neutrophil extracellular traps kill bacteria Science 2004303(5663):1532–1535. - PubMed
-
- Engelmann B, Massberg S. Thrombosis as an intravascular effector of innate immunity. Nat Rev Immunol. 2013;13(01):34–45. - PubMed
-
- Gould T J, Lysov Z, Liaw P C. Extracellular DNA and histones: double-edged swords in immunothrombosis. J Thromb Haemost. 2015;13 01:S82–S91. - PubMed