

Ecological limits to evolutionary rescue
- PMID: 33131439
- PMCID: PMC7662203
- DOI: 10.1098/rstb.2019.0453
Ecological limits to evolutionary rescue
Abstract
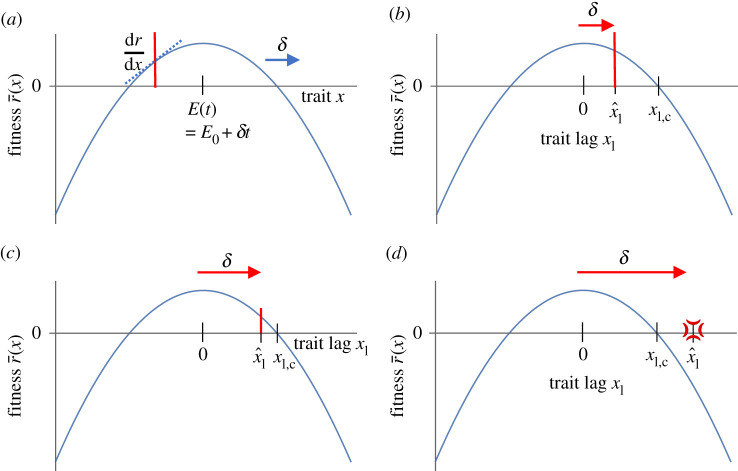
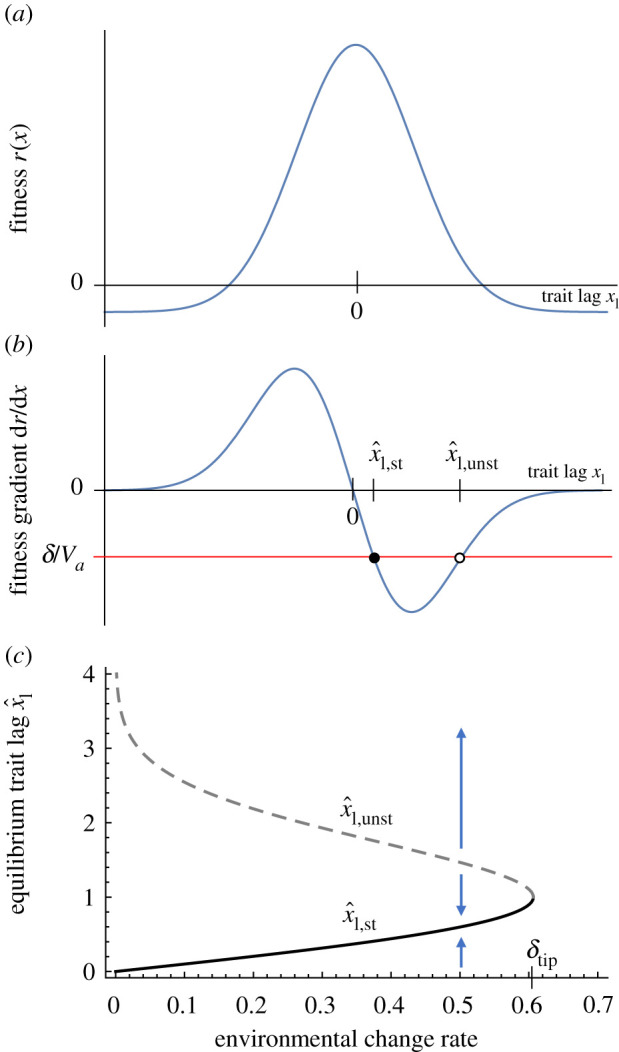
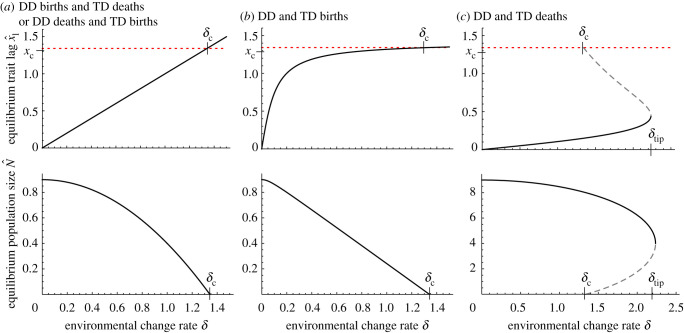
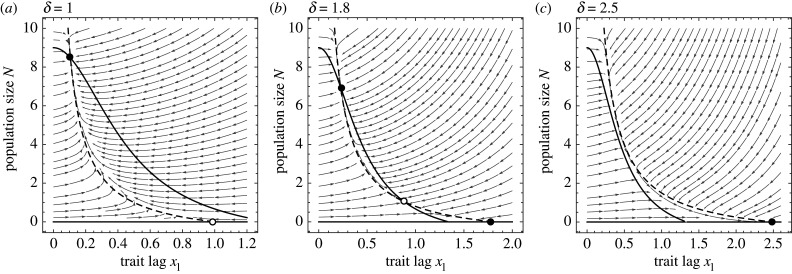
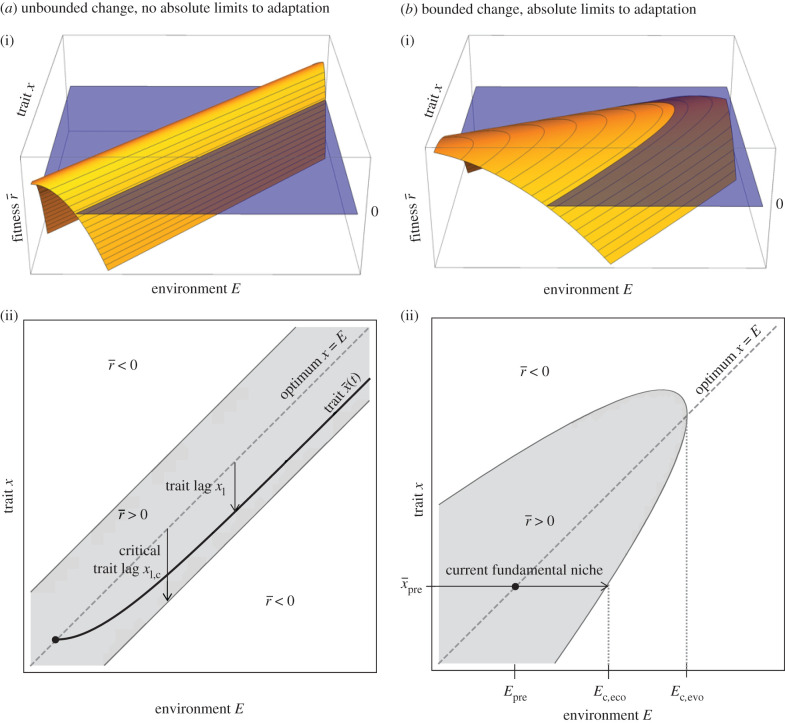
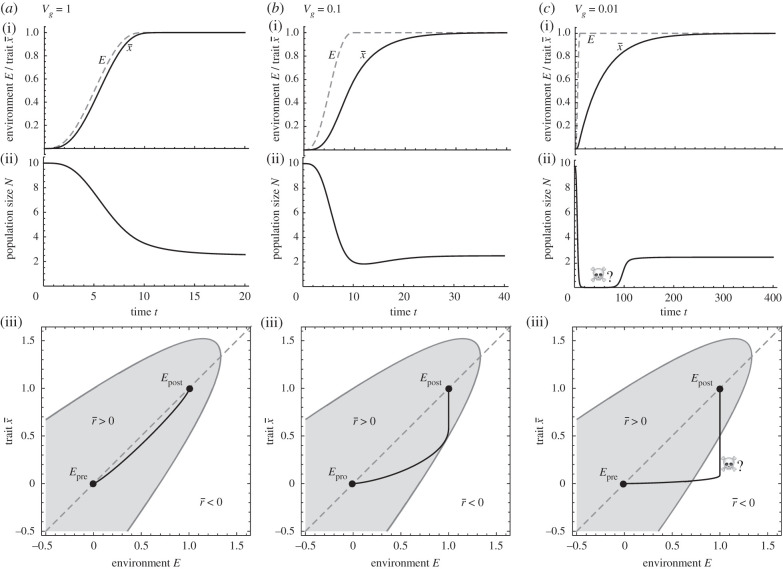
Environments change, for both natural and anthropogenic reasons, which can threaten species persistence. Evolutionary adaptation is a potentially powerful mechanism to allow species to persist in these changing environments. To determine the conditions under which adaptation will prevent extinction (evolutionary rescue), classic quantitative genetics models have assumed a constantly changing environment. They predict that species traits will track a moving environmental optimum with a lag that approaches a constant. If fitness is negative at this lag, the species will go extinct. There have been many elaborations of these models incorporating increased genetic realism. Here, we review and explore the consequences of four ecological complications: non-quadratic fitness functions, interacting density- and trait-dependence, species interactions and fundamental limits to adaptation. We show that non-quadratic fitness functions can result in evolutionary tipping points and existential crises, as can the interaction between density- and trait-dependent mortality. We then review the literature on how interspecific interactions affect adaptation and persistence. Finally, we suggest an alternative theoretical framework that considers bounded environmental change and fundamental limits to adaptation. A research programme that combines theory and experiments and integrates across organizational scales will be needed to predict whether adaptation will prevent species extinction in changing environments. This article is part of the theme issue 'Integrative research perspectives on marine conservation'.
Keywords: climate change; eco-evolutionary dynamics; environmental change; evolutionary rescue; moving optimum; quantitative genetics.
Conflict of interest statement
We declare we have no competing interests.
Figures
References
-
- Bell G. 2017. Evolutionary rescue. Annu. Rev. Ecol. Evol. Syst. 48, 605–627. (10.1146/annurev-ecolsys-110316-023011) - DOI
Publication types
MeSH terms
Associated data
LinkOut - more resources
Full Text Sources