

CEACAMs serve as toxin-stimulated receptors for enterotoxigenic Escherichia coli
- PMID: 33139570
- PMCID: PMC7682567
- DOI: 10.1073/pnas.2012480117
CEACAMs serve as toxin-stimulated receptors for enterotoxigenic Escherichia coli
Abstract
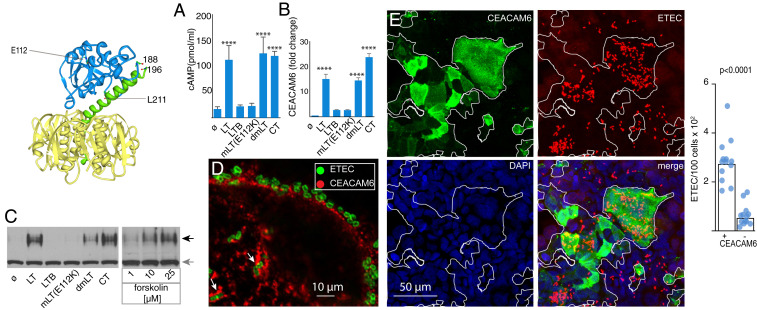
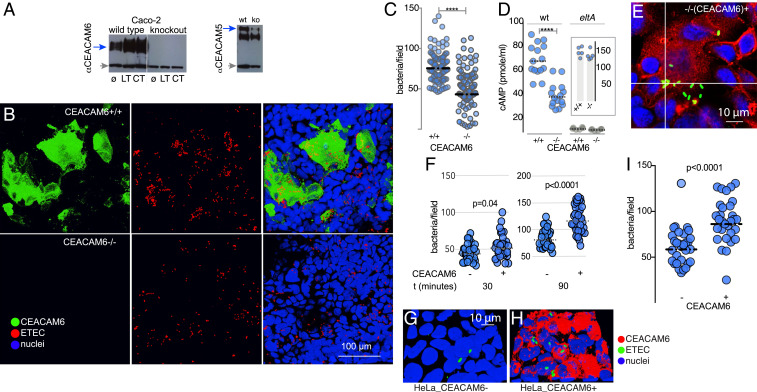
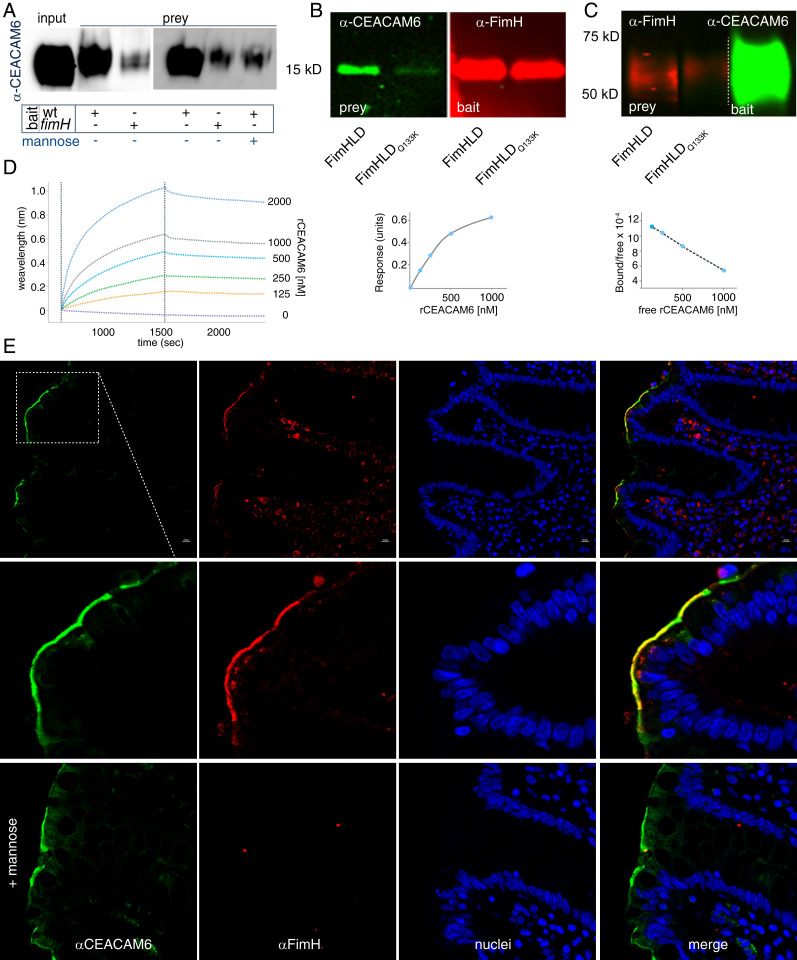
The enterotoxigenic Escherichia coli (ETEC) are among the most common causes of diarrheal illness and death due to diarrhea among young children in low-/middle-income countries (LMICs). ETEC have also been associated with important sequelae including malnutrition and stunting, placing children at further risk of death from diarrhea and other infections. Our understanding of the molecular pathogenesis of acute diarrheal disease as well as the sequelae linked to ETEC are still evolving. It has long been known that ETEC heat-labile toxin (LT) activates production of cAMP in the cell, signaling the modulation of cellular ion channels that results in a net efflux of salt and water into the intestinal lumen, culminating in watery diarrhea. However, as LT also promotes ETEC adhesion to intestinal epithelial cells, we postulated that increases in cAMP, a critical cellular "second messenger," may be linked to changes in cellular architecture that favor pathogen-host interactions. Indeed, here we show that ETEC use LT to up-regulate carcinoembryonic antigenrelated cell adhesion molecules (CEACAMs) on the surface of small intestinal epithelia, where they serve as critical bacterial receptors. Moreover, we show that bacteria are specifically recruited to areas of CEACAM expression, in particular CEACAM6, and that deletion of this CEACAM abrogates both bacterial adhesion and toxin delivery. Collectively, these results provide a paradigm for the molecular pathogenesis of ETEC in which the bacteria use toxin to drive up-regulation of cellular targets that enhances subsequent pathogen-host interactions.
Keywords: CEACAM; diarrhea; enterotoxin.
Conflict of interest statement
The authors declare no competing interest.
Figures

References
-
- MAL-ED Network Investigators , The MAL-ED study: A multinational and multidisciplinary approach to understand the relationship between enteric pathogens, malnutrition, gut physiology, physical growth, cognitive development, and immune responses in infants and children up to 2 years of age in resource-poor environments. Clin. Infect. Dis. 59 (suppl. 4), S193–S206 (2014). - PubMed
Publication types
MeSH terms
Substances
Associated data
- figshare/10.6084/m9.figshare.12137544
- figshare/10.6084/m9.figshare.12141033
- figshare/10.6084/m9.figshare.121412-28
- figshare/10.6084/m9.figshare.12137658
- figshare/10.6084/m9.figshare.12137652
- figshare/10.6084/m9.figshare.12137655
- figshare/10.6084/m9.figshare.12141138
- figshare/10.6084/m9.figshare.12137661
- figshare/10.6084/m9.figshare.12138297
- figshare/10.6084/m9.figshare.12142092
- figshare/10.6084/m9.figshare.12140994
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
