

Gephyrin-mediated formation of inhibitory postsynaptic density sheet via phase separation
- PMID: 33139925
- PMCID: PMC8027005
- DOI: 10.1038/s41422-020-00433-1
Gephyrin-mediated formation of inhibitory postsynaptic density sheet via phase separation
Abstract
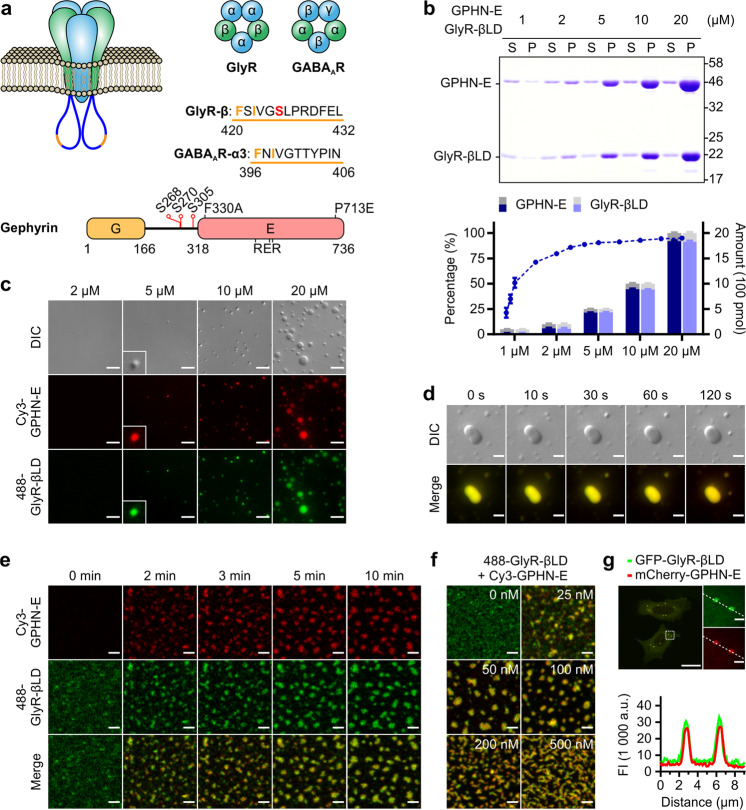
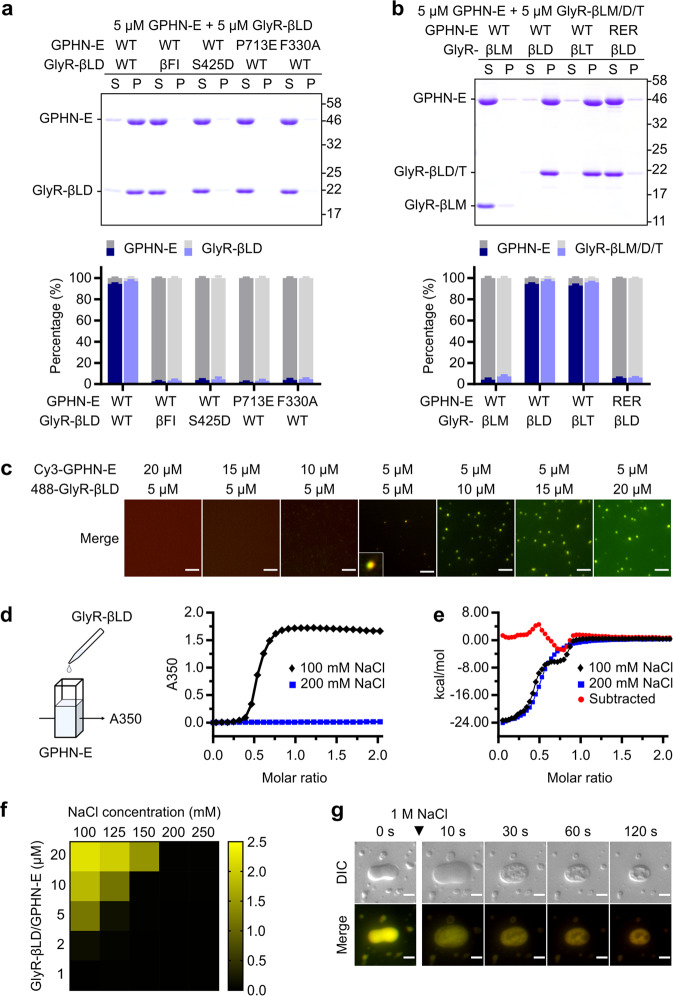
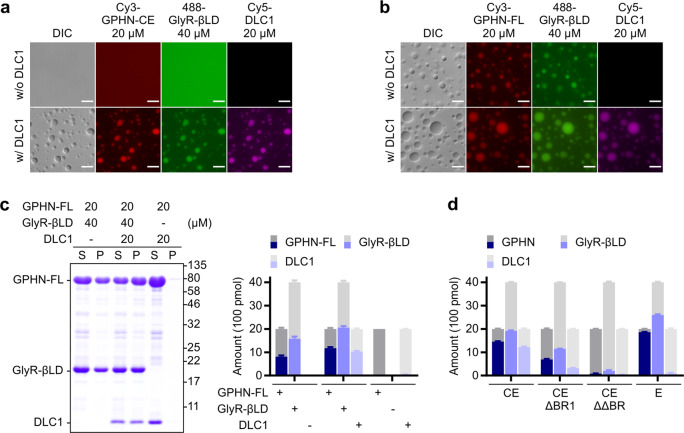
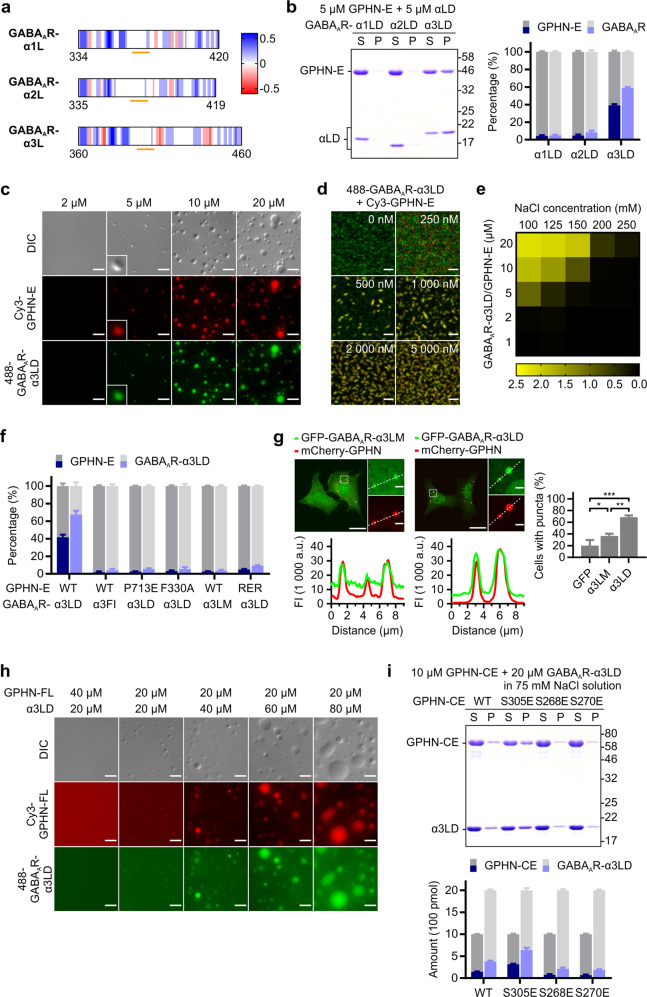
Inhibitory synapses are also known as symmetric synapses due to their lack of prominent postsynaptic densities (PSDs) under a conventional electron microscope (EM). Recent cryo-EM tomography studies indicated that inhibitory synapses also contain PSDs, albeit with a rather thin sheet-like structure. It is not known how such inhibitory PSD (iPSD) sheet might form. Here, we demonstrate that the key inhibitory synapse scaffold protein gephyrin, when in complex with either glycine or GABAA receptors, spontaneously forms highly condensed molecular assemblies via phase separation both in solution and on supported membrane bilayers. Multivalent and specific interactions between the dimeric E-domain of gephyrin and the glycine/GABAA receptor multimer are essential for the iPSD condensate formation. Gephyrin alone does not form condensates. The linker between the G- and E-domains of gephyrin inhibits the iPSD condensate formation via autoinhibition. Phosphorylation of specific residues in the linker or binding of target proteins such as dynein light chain to the linker domain regulates gephyrin-mediated glycine/GABAA receptor clustering. Thus, analogous to excitatory PSDs, iPSDs are also formed by phase separation-mediated condensation of scaffold protein/neurotransmitter receptor complexes.
Conflict of interest statement
The authors declare no competing interests.
Figures



Comment in
-
Gephyrin: a scaffold that builds a phase at the inhibitory postsynapses.Cell Res. 2021 Mar;31(3):245-246. doi: 10.1038/s41422-020-00440-2. Cell Res. 2021. PMID: 33203920 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
