

Metabolic reprogramming of glycolysis and glutamine metabolism are key events in myofibroblast transition in systemic sclerosis pathogenesis
- PMID: 33140521
- PMCID: PMC7754020
- DOI: 10.1111/jcmm.16013
Metabolic reprogramming of glycolysis and glutamine metabolism are key events in myofibroblast transition in systemic sclerosis pathogenesis
Abstract
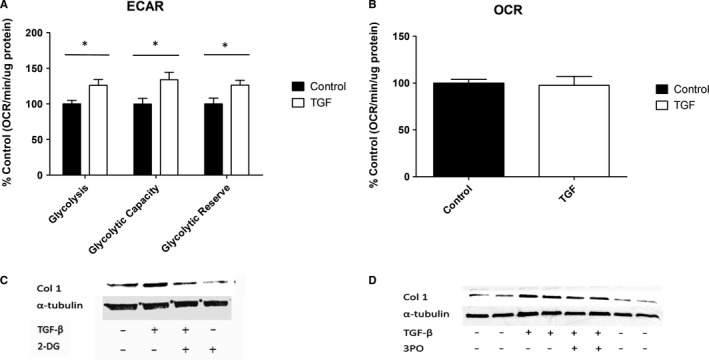
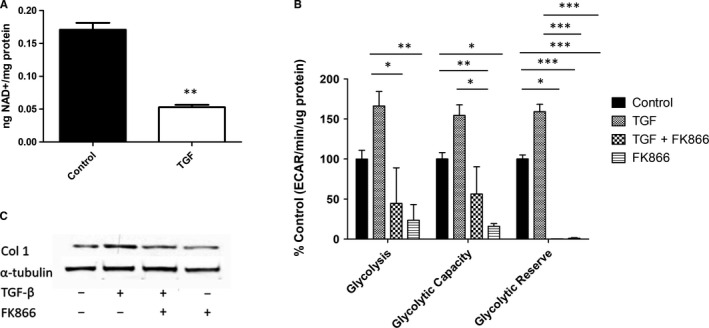
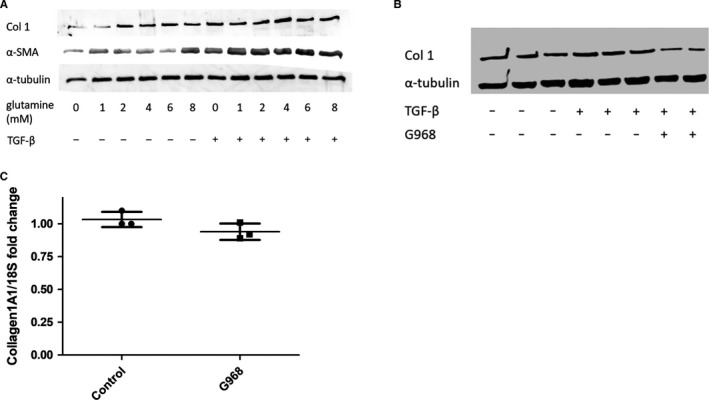
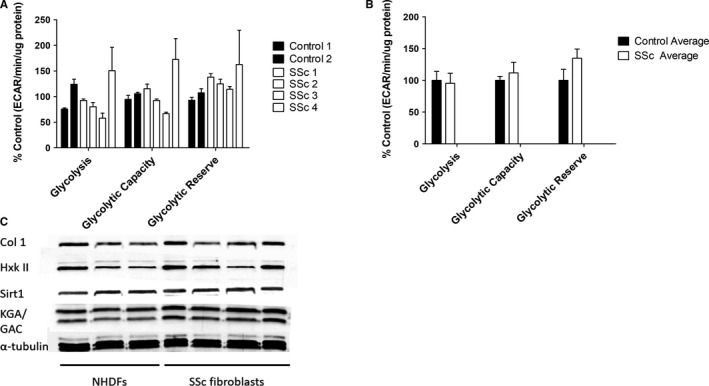
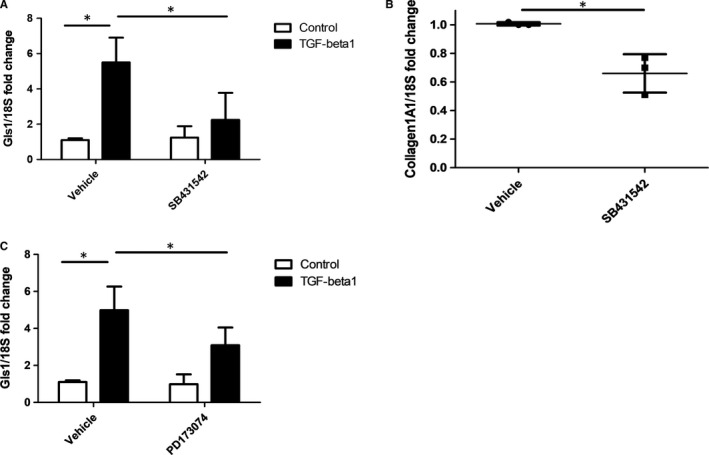
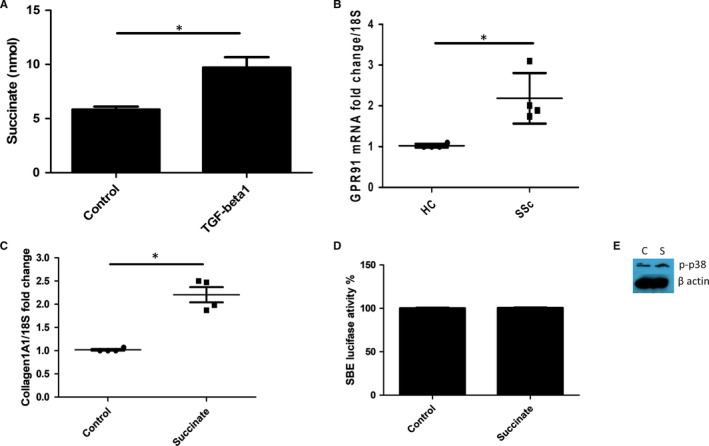
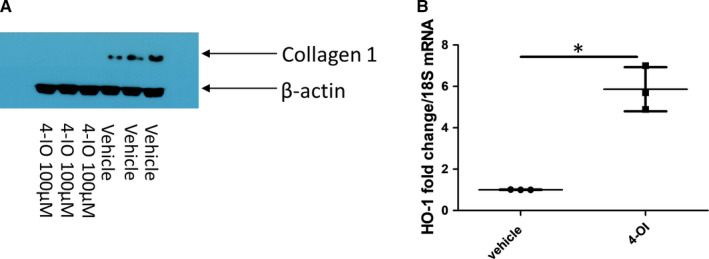
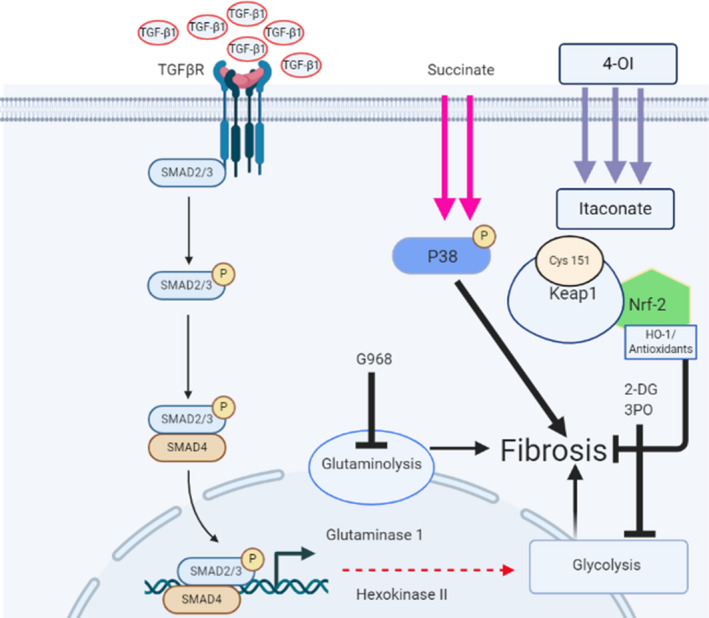
Systemic Sclerosis (SSc) is a rare fibrotic autoimmune disorder for which no curative treatments currently exist. Metabolic remodelling has recently been implicated in other autoimmune diseases; however, its potential role in SSc has received little attention. Here, we aimed to determine whether changes to glycolysis and glutaminolysis are important features of skin fibrosis. TGF-β1, the quintessential pro-fibrotic stimulus, was used to activate fibrotic pathways in NHDFs in vitro. Dermal fibroblasts derived from lesions of SSc patients were also used for in vitro experiments. Parameters of glycolytic function were assessed using by measuring extracellular acidification in response to glycolytic activators/inhibitors, whilst markers of fibrosis were measured by Western blotting following the use of the glycolysis inhibitors 2-dg and 3PO and the glutaminolysis inhibitor G968. Succinate was also measured after TGF-β1 stimulation. Itaconate was added to SSc fibroblasts and collagen examined. TGF-β1 up-regulates glycolysis in dermal fibroblasts, and inhibition of glycolysis attenuates its pro-fibrotic effects. Furthermore, inhibition of glutamine metabolism also reverses TGF-β1-induced fibrosis, whilst glutaminase expression is up-regulated in dermal fibroblasts derived from SSc patient skin lesions, suggesting that enhanced glutamine metabolism is another aspect of the pro-fibrotic metabolic phenotype in skin fibrosis. TGF-β1 was also able to enhance succinate production, with increased succinate shown to be associated with increased collagen expression. Incubation of SSc cells with itaconate, an important metabolite, reduced collagen expression. TGF-β1 activation of glycolysis is a key feature of the fibrotic phenotype induced by TGF-B1 in skin cells, whilst increased glutaminolysis is also evident in SSc fibroblasts.
Keywords: Systemic Sclerosis; TGF-β1; fibrosis; glutaminolysis; glycolysis.
© 2020 The Authors. Journal of Cellular and Molecular Medicine published by Foundation for Cellular and Molecular Medicine and John Wiley & Sons Ltd.
Conflict of interest statement
The authors state there are no conflicts of interest to declare.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
