

Regulation of peroxisomal trafficking and distribution
- PMID: 33141311
- PMCID: PMC7966214
- DOI: 10.1007/s00018-020-03687-5
Regulation of peroxisomal trafficking and distribution
Abstract
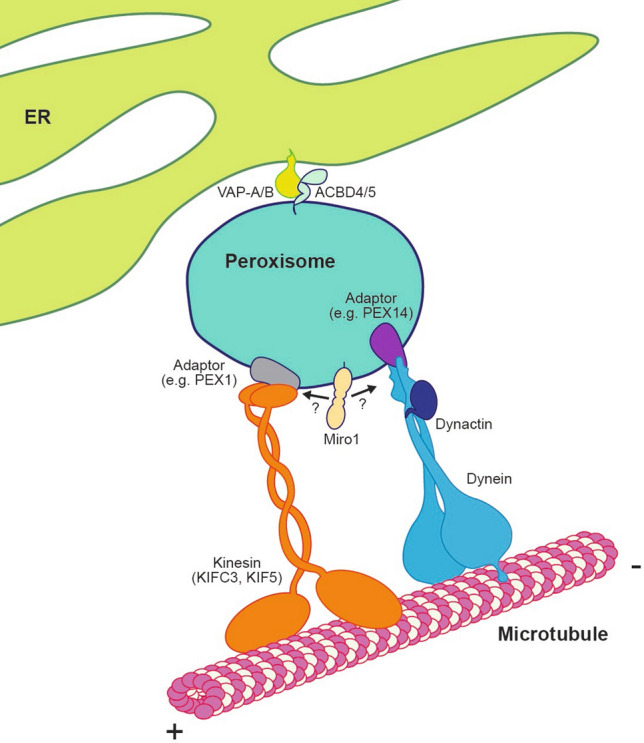
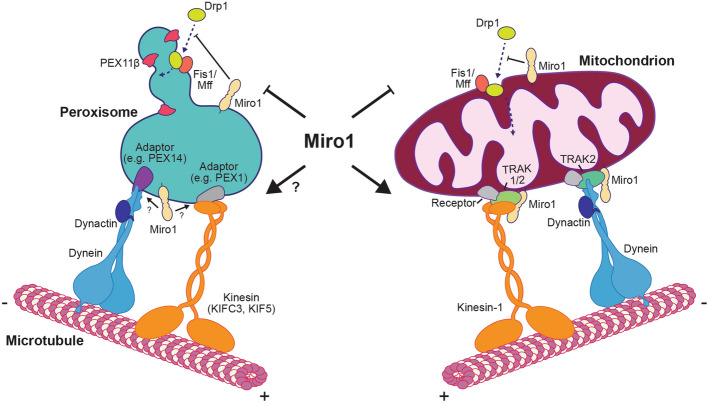
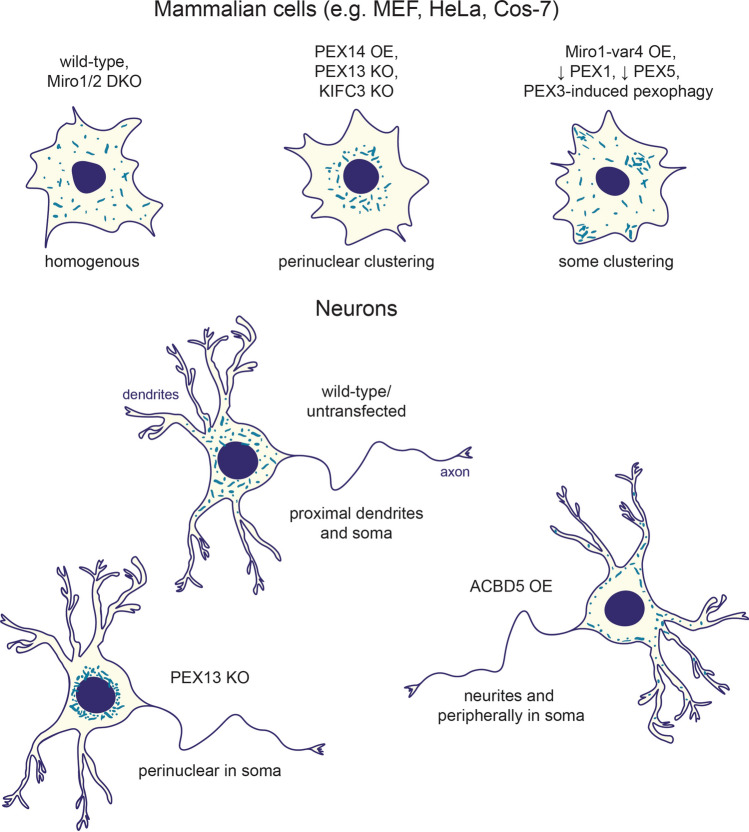
Peroxisomes are organelles that perform a wide range of essential metabolic processes. To ensure that peroxisomes are optimally positioned in the cell, they must be transported by both long- and short-range trafficking events in response to cellular needs. Here, we review our current understanding of the mechanisms by which the cytoskeleton and organelle contact sites alter peroxisomal distribution. Though the focus of the review is peroxisomal transport in mammalian cells, findings from flies and fungi are used for comparison and to inform the gaps in our understanding. Attention is given to the apparent overlap in regulatory mechanisms for mitochondrial and peroxisomal trafficking, along with the recently discovered role of the mitochondrial Rho-GTPases, Miro, in peroxisomal dynamics. Moreover, we outline and discuss the known pathological and pharmacological conditions that perturb peroxisomal positioning. We conclude by highlighting several gaps in our current knowledge and suggest future directions that require attention.
Keywords: Actin; Disease; Dynein; Kinesin; Microtubule.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
