

Technical advance: The use of tree shrews as a model of pulmonary fibrosis
- PMID: 33141839
- PMCID: PMC7608928
- DOI: 10.1371/journal.pone.0241323
Technical advance: The use of tree shrews as a model of pulmonary fibrosis
Abstract
Background: Idiopathic pulmonary fibrosis (IPF) is a chronic, progressive disease with a high morbidity and mortality. Some of the mechanisms of fibrosis development have been described using rodent models; however, the relevance of findings in these animal models is difficult to assess. New innovative models are needed that closely mimic IPF disease pathology.
Methods: To overcome this unmet need of investigating IPF with a relevant model, we utilized tree shrews, which are genetically, anatomically, and metabolically similar to primates and humans. Using human antibodies and primers, we investigated the role of macrophage phenotypic switching in normal and IPF subjects and bleomycin-injured tree shrews.
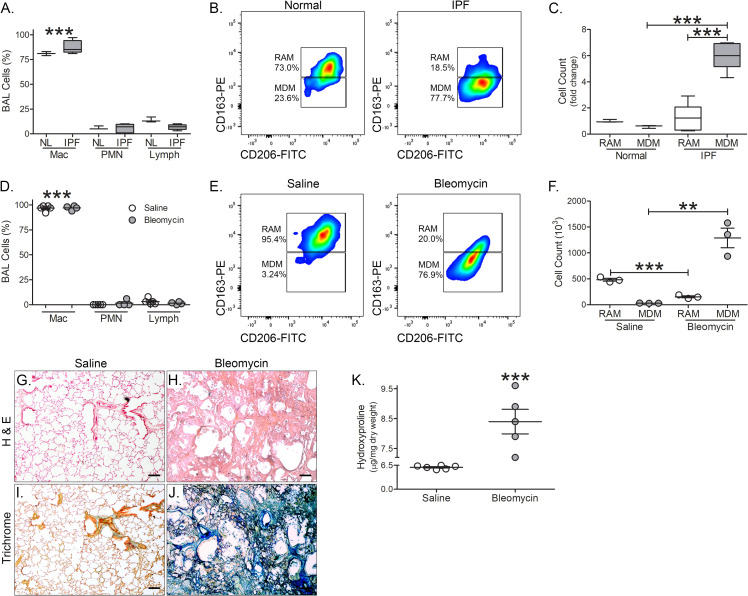
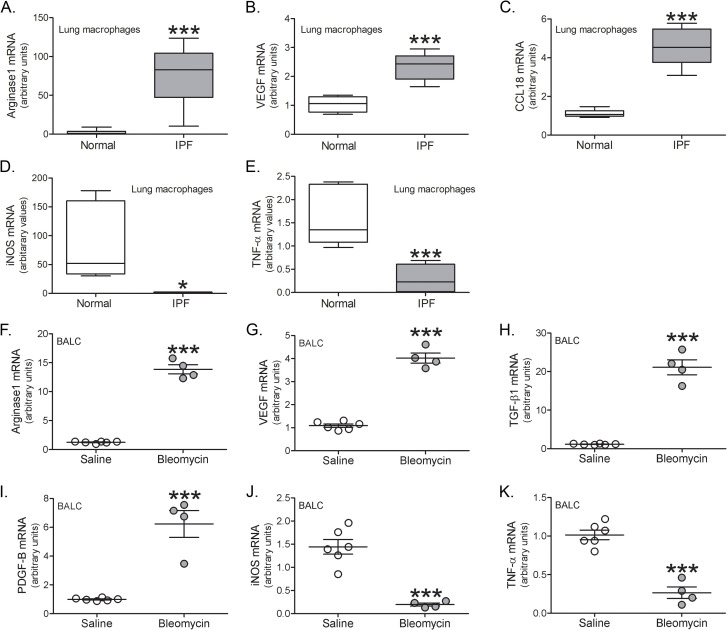
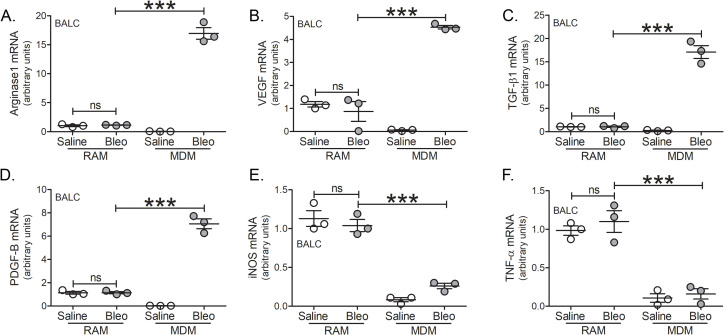
Results: Bronchoalveolar lavage (BAL) cells from tree shrews expressed human markers, and there was recruitment of monocyte-derived macrophages (MDMs) to the lung in IPF subjects and bleomycin-injured tree shrews. MDMs were polarized to a profibrotic phenotype in IPF and in bleomycin-injured tree shrews. Resident alveolar macrophages (RAMs) expressed proinflammatory markers regardless of bleomycin exposure. Tree shrews developed bleomycin-induced pulmonary fibrosis with architectural distortion in parenchyma and widespread collagen deposition.
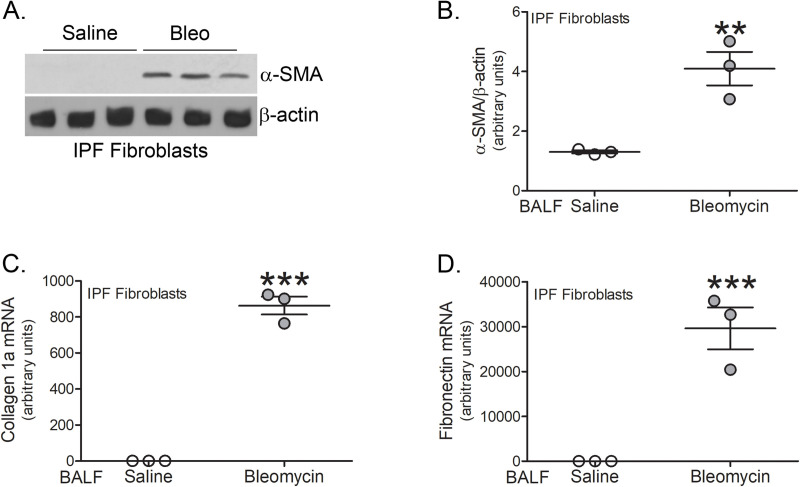
Conclusion: The profibrotic polarization of macrophages has been demonstrated to be present in IPF subjects and in fibrotic mice. Although the lung macrophages have long been considered to be homogeneous, recent evidence indicates that these cells are heterogeneous during multiple chronic lung diseases. Here, we show new data that indicate a critical and essential role for macrophage-fibroblast crosstalk promoting fibroblast differentiation and collagen production. in the development and progression of fibrosis. The current data strongly suggest development of therapeutics that attenuate of the profibrotic activation of MDMs may mitigate macrophage-fibroblast interaction. These observations demonstrate that tree shrews are an ideal animal model to investigate the pathogenesis of IPF as they are genetically, anatomically, and metabolically closer to humans than the more commonly used rodent models.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
A novel tree shrew model of pulmonary fibrosis.Lab Invest. 2021 Jan;101(1):116-124. doi: 10.1038/s41374-020-00476-3. Epub 2020 Aug 9. Lab Invest. 2021. PMID: 32773774
-
Single-Cell RNA Sequencing Reveals Monocyte-Derived Interstitial Macrophages with a Pro-Fibrotic Phenotype in Bleomycin-Induced Pulmonary Fibrosis.Int J Mol Sci. 2024 Oct 30;25(21):11669. doi: 10.3390/ijms252111669. Int J Mol Sci. 2024. PMID: 39519222 Free PMC article.
-
CD148 Deficiency in Fibroblasts Promotes the Development of Pulmonary Fibrosis.Am J Respir Crit Care Med. 2021 Aug 1;204(3):312-325. doi: 10.1164/rccm.202008-3100OC. Am J Respir Crit Care Med. 2021. PMID: 33784491 Free PMC article.
-
The pathogenesis of bleomycin-induced lung injury in animals and its applicability to human idiopathic pulmonary fibrosis.Exp Lung Res. 2015 Mar;41(2):57-73. doi: 10.3109/01902148.2014.979516. Epub 2014 Dec 16. Exp Lung Res. 2015. PMID: 25514507 Review.
-
Allies or enemies? The effect of regulatory T cells and related T lymphocytes on the profibrotic environment in bleomycin-injured lung mouse models.Clin Exp Med. 2023 Aug;23(4):1075-1088. doi: 10.1007/s10238-022-00945-7. Epub 2022 Nov 20. Clin Exp Med. 2023. PMID: 36403186 Free PMC article. Review.
Cited by
-
Myeloid Heterogeneity Mediates Acute Exacerbations of Pulmonary Fibrosis.J Immunol. 2023 Dec 1;211(11):1714-1724. doi: 10.4049/jimmunol.2300053. J Immunol. 2023. PMID: 37782053 Free PMC article.
-
Cell Adhesion Molecules in Fibrotic Diseases.Biomedicines. 2023 Jul 14;11(7):1995. doi: 10.3390/biomedicines11071995. Biomedicines. 2023. PMID: 37509634 Free PMC article. Review.
-
The Mongolian gerbil as an advanced model to study cone system physiology.Front Cell Neurosci. 2024 Jan 25;18:1339282. doi: 10.3389/fncel.2024.1339282. eCollection 2024. Front Cell Neurosci. 2024. PMID: 38333056 Free PMC article.
-
Communication between alveolar macrophages and fibroblasts via the TNFSF12-TNFRSF12A pathway promotes pulmonary fibrosis in severe COVID-19 patients.J Transl Med. 2024 Jul 29;22(1):698. doi: 10.1186/s12967-024-05381-7. J Transl Med. 2024. PMID: 39075394 Free PMC article.
-
Study of tree shrew biology and models: A booming and prosperous field for biomedical research.Zool Res. 2024 Jul 18;45(4):877-909. doi: 10.24272/j.issn.2095-8137.2024.199. Zool Res. 2024. PMID: 39004865 Free PMC article. Review.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
