

Neurons Release Serine to Support mRNA Translation in Pancreatic Cancer
- PMID: 33142117
- PMCID: PMC8100789
- DOI: 10.1016/j.cell.2020.10.016
Neurons Release Serine to Support mRNA Translation in Pancreatic Cancer
Abstract
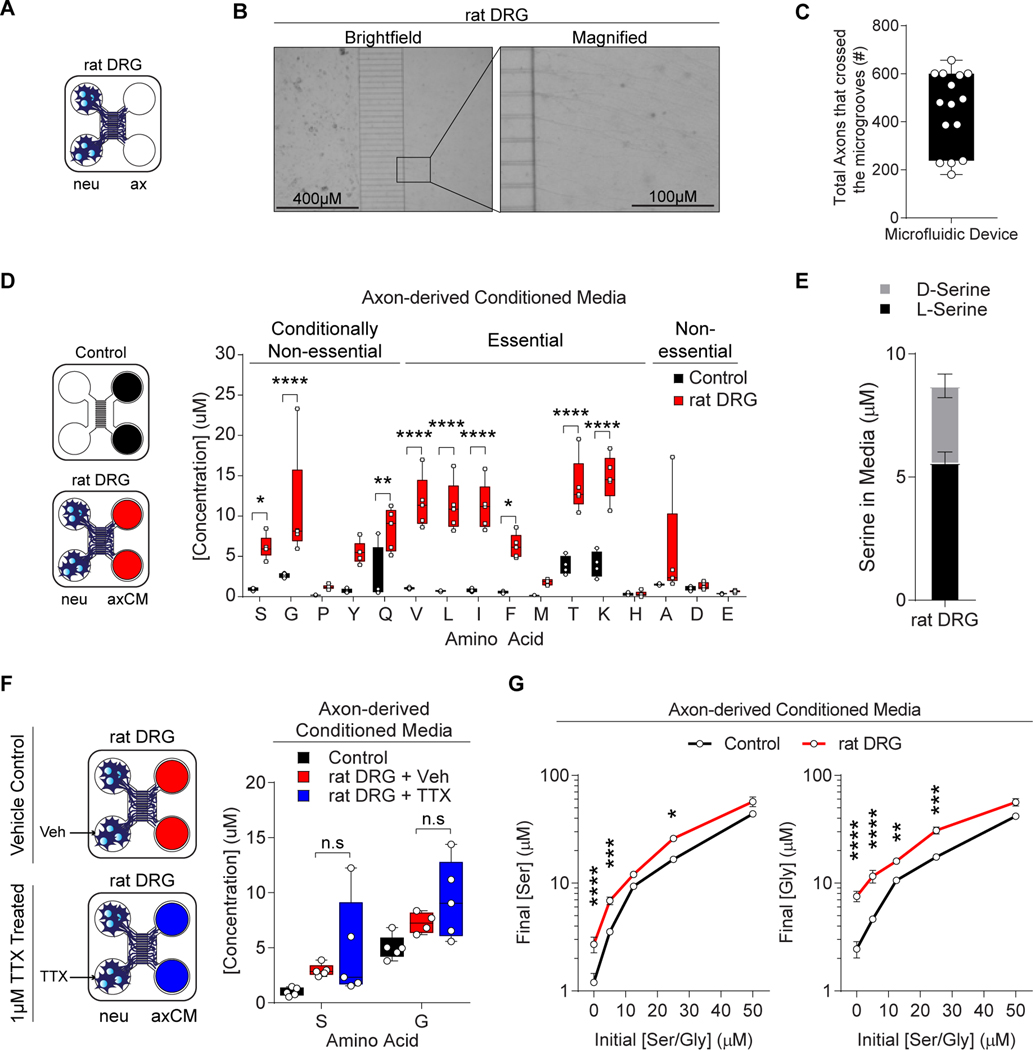
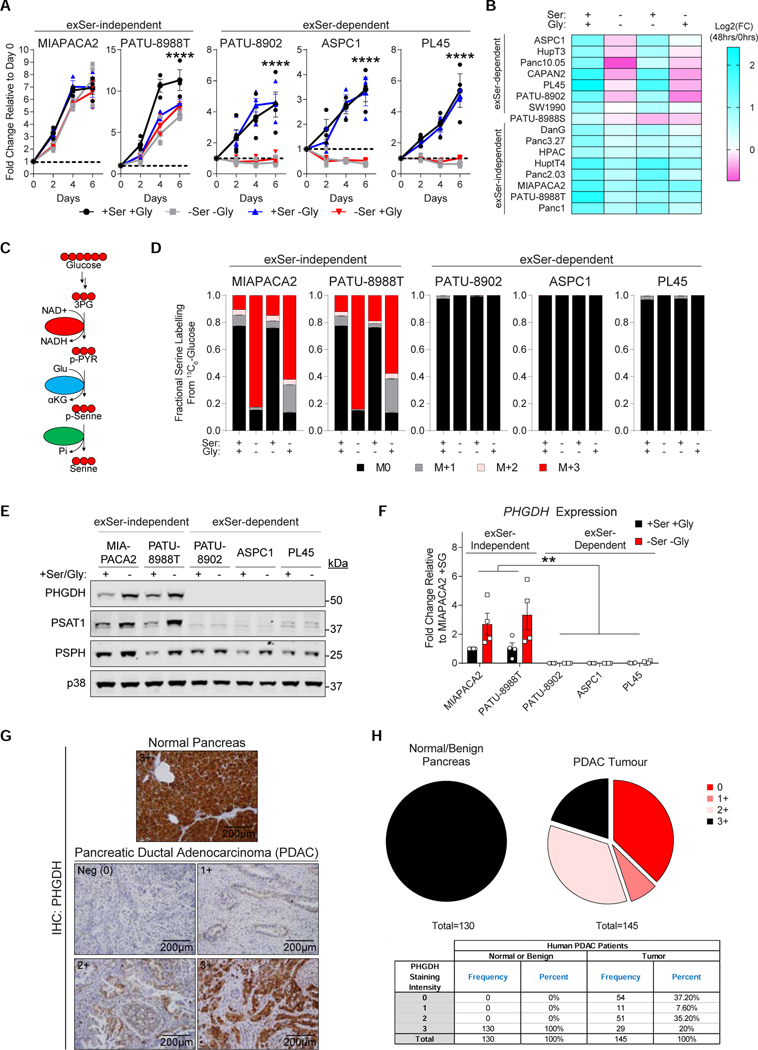
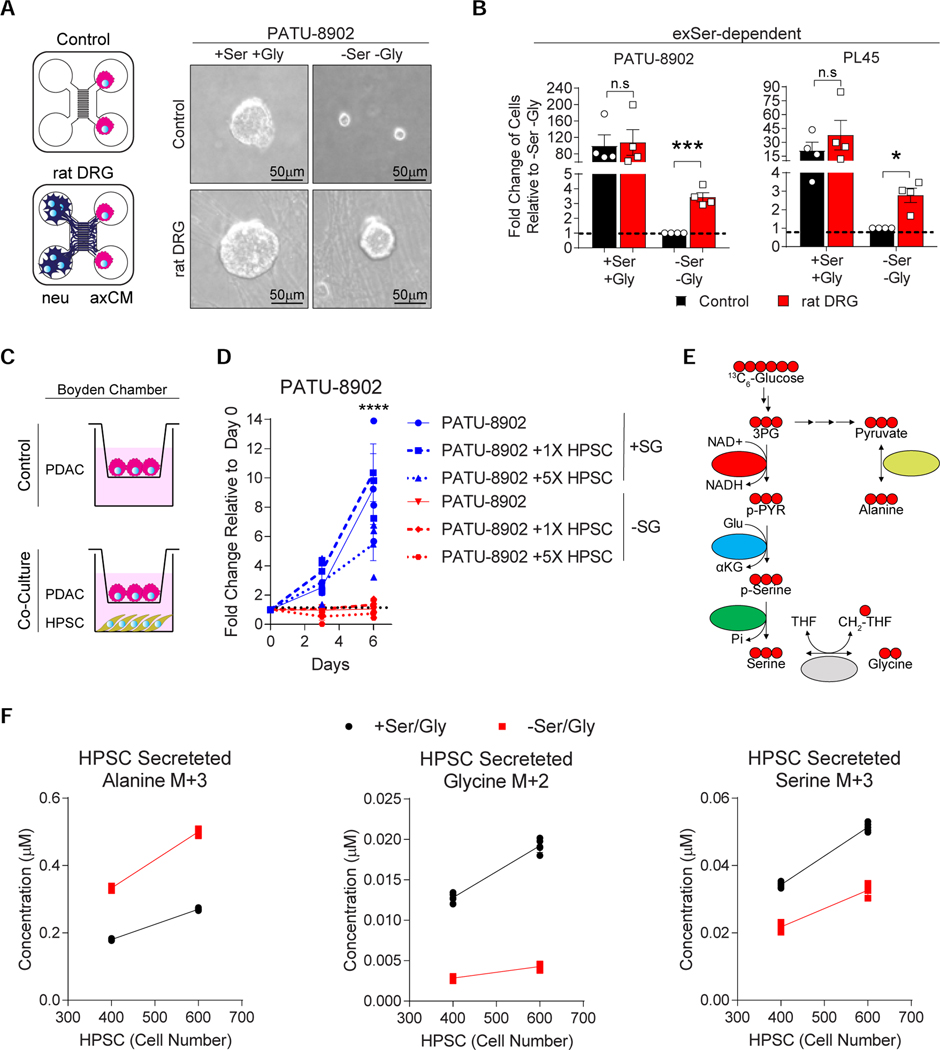
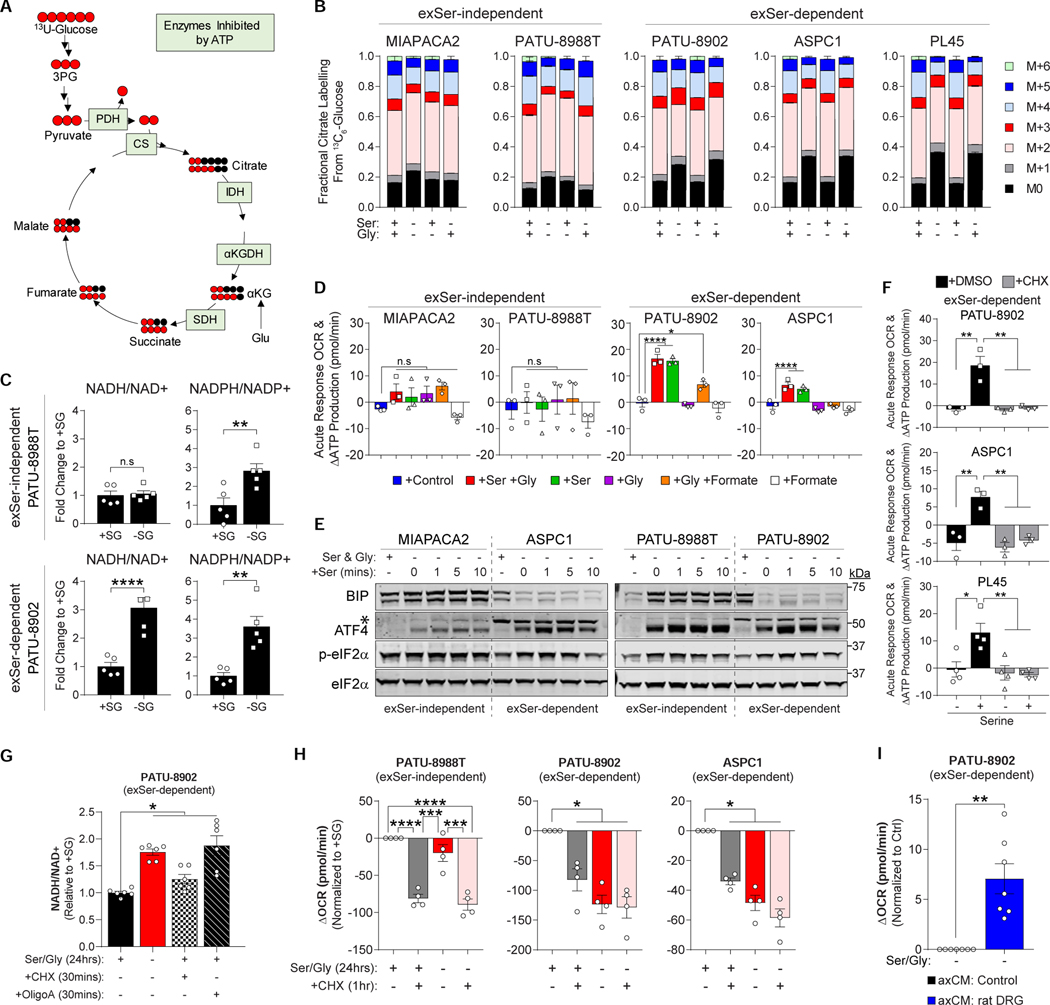
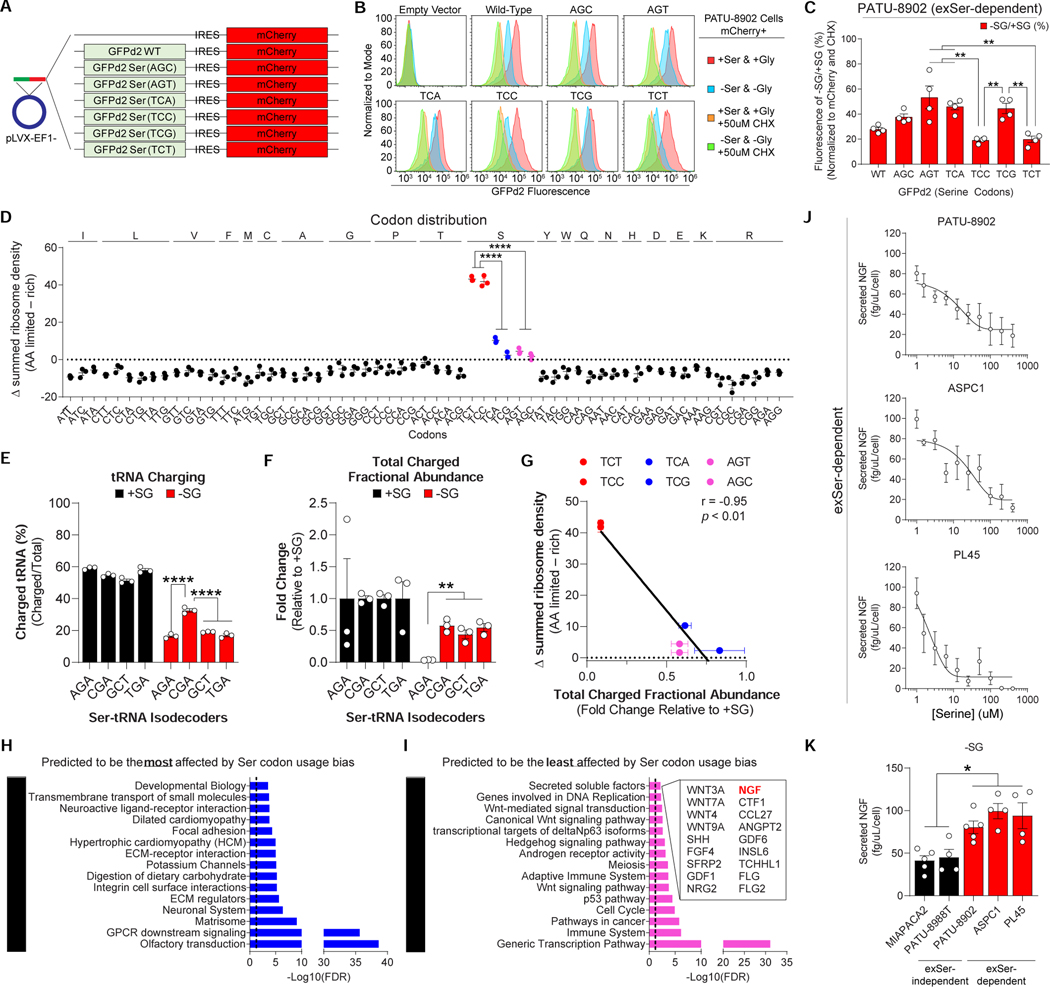
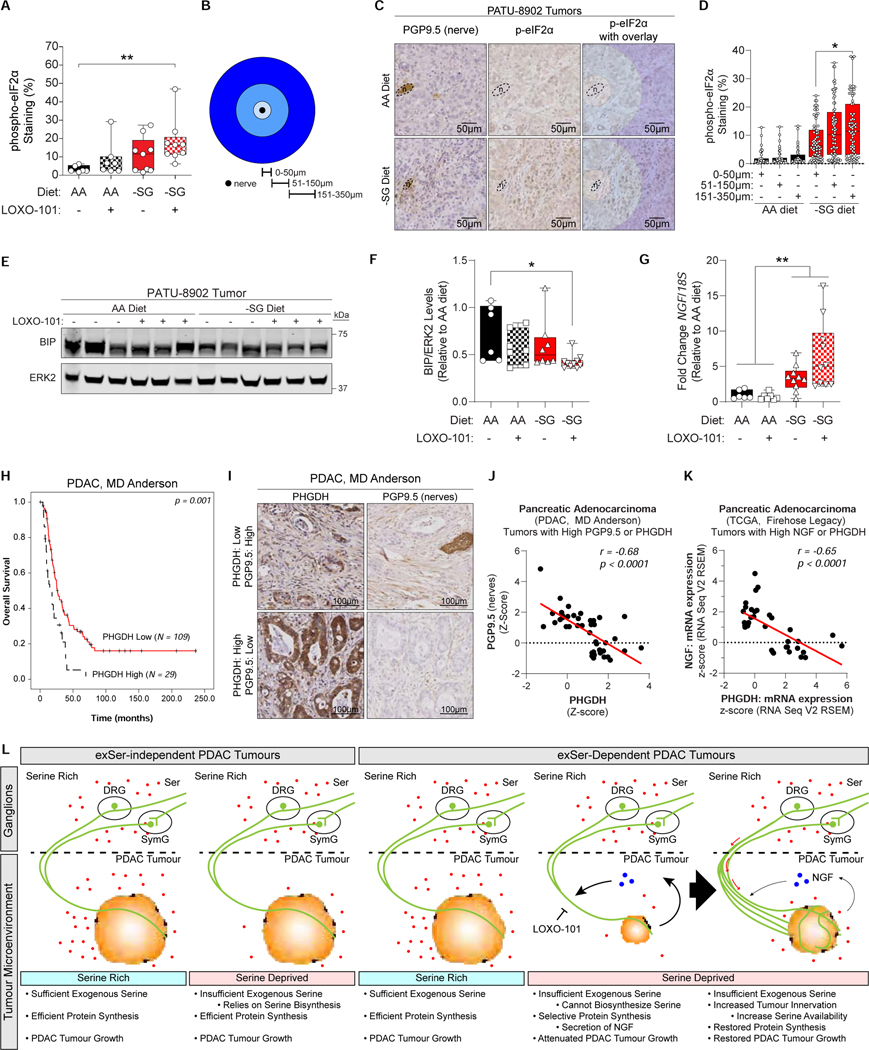
Pancreatic ductal adenocarcinoma (PDAC) tumors have a nutrient-poor, desmoplastic, and highly innervated tumor microenvironment. Although neurons can release stimulatory factors to accelerate PDAC tumorigenesis, the metabolic contribution of peripheral axons has not been explored. We found that peripheral axons release serine (Ser) to support the growth of exogenous Ser (exSer)-dependent PDAC cells during Ser/Gly (glycine) deprivation. Ser deprivation resulted in ribosomal stalling on two of the six Ser codons, TCC and TCT, and allowed the selective translation and secretion of nerve growth factor (NGF) by PDAC cells to promote tumor innervation. Consistent with this, exSer-dependent PDAC tumors grew slower and displayed enhanced innervation in mice on a Ser/Gly-free diet. Blockade of compensatory neuronal innervation using LOXO-101, a Trk-NGF inhibitor, further decreased PDAC tumor growth. Our data indicate that axonal-cancer metabolic crosstalk is a critical adaptation to support PDAC growth in nutrient poor environments.
Keywords: mRNA translation; metabolic crosstalk; neurons; pancreatic cancer; serine.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests M.E.P. has options in Raze Therapeutics and received travel funds from Thermo Fisher Scientific. J.D.M is an inventor on a patent pertaining to the autophagic control of iron metabolism. A.C.K. has financial interests in Vescor Therapeutics, LLC. A.C.K. is an inventor on patents pertaining to KRAS regulated metabolic pathways, redox control pathways in pancreatic cancer, targeting GOT1 as a therapeutic approach, and the autophagic control of iron metabolism. A.C.K is on the Science Advisory Board of Rafael/Cornerstone Pharma. A.C.K. has been a consultant for Deciphera Pharma. The other authors declare no competing interest.
Figures

Comment in
-
Recruited Nerves Supply Serine to Support Pancreatic Cancer Growth.Cancer Discov. 2021 Jan;11(1):11. doi: 10.1158/2159-8290.CD-RW2020-165. Epub 2020 Nov 13. Cancer Discov. 2021. PMID: 33188053
-
Neuronal innervation supports PDAC growth via release of serine.Nat Rev Gastroenterol Hepatol. 2021 Jan;18(1):5. doi: 10.1038/s41575-020-00394-1. Nat Rev Gastroenterol Hepatol. 2021. PMID: 33257832 No abstract available.
-
The neuronal-metabolic interface.Nat Rev Cancer. 2021 Feb;21(2):68. doi: 10.1038/s41568-020-00324-y. Nat Rev Cancer. 2021. PMID: 33262455 No abstract available.
-
Nerves on tr[ac]k to support pancreatic cancer metabolism.Cell Res. 2021 Apr;31(4):381-382. doi: 10.1038/s41422-020-00462-w. Cell Res. 2021. PMID: 33536521 Free PMC article. No abstract available.
References
-
- Abbaci M, Barberi-Heyob M, Blondel W, Guillemin F, and Didelon J. (2008). Advantages and limitations of commonly used methods to assay the molecular permeability of gap junctional intercellular communication. BioTechniques 45, 33–62. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
