

Expression of EMT-Related Genes in Hybrid E/M Colorectal Cancer Cells Determines Fibroblast Activation and Collagen Remodeling
- PMID: 33143259
- PMCID: PMC7662237
- DOI: 10.3390/ijms21218119
Expression of EMT-Related Genes in Hybrid E/M Colorectal Cancer Cells Determines Fibroblast Activation and Collagen Remodeling
Abstract
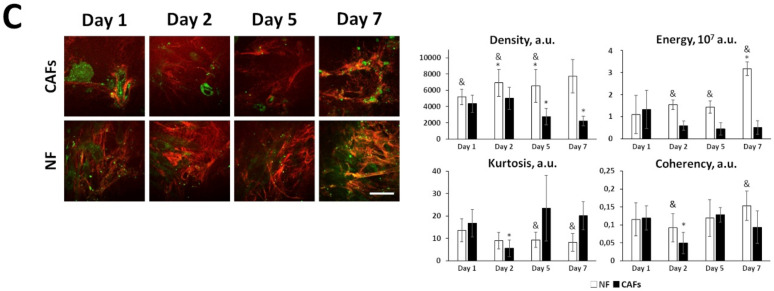
Collagen, the main non-cellular component of the extracellular matrix (ECM), is profoundly reorganized during tumorigenesis and has a strong impact on tumor behavior. The main source of collagen in tumors is cancer-associated fibroblasts. Cancer cells can also participate in the synthesis of ECM; however, the contribution of both types of cells to collagen rearrangements during the tumor progression is far from being clear. Here, we investigated the processes of collagen biosynthesis and remodeling in parallel with the transcriptome changes during cancer cells and fibroblasts interactions. Combining immunofluorescence, RNA sequencing, and second harmonic generation microscopy, we have explored the relationships between the ratio of epithelial (E) and mesenchymal (M) components of hybrid E/M cancer cells, their ability to activate fibroblasts, and the contributions of both cell types to collagen remodeling. To this end, we studied (i) co-cultures of colorectal cancer cells and normal fibroblasts in a collagen matrix, (ii) patient-derived cancer-associated fibroblasts, and (iii) mouse xenograft models. We found that the activation of normal fibroblasts that form dense collagen networks consisting of large, highly oriented fibers depends on the difference in E/M ratio in the cancer cells. The more-epithelial cells activate the fibroblasts more strongly, which correlates with a dense and highly ordered collagen structure in tumors in vivo. The more-mesenchymal cells activate the fibroblasts to a lesser degree; on the other hand, this cell line has a higher innate collagen remodeling capacity. Normal fibroblasts activated by cancer cells contribute to the organization of the extracellular matrix in a way that is favorable for migratory potency. At the same time, in co-culture with epithelial cancer cells, the contribution of fibroblasts to the reorganization of ECM is more pronounced. Therefore, one can expect that targeting the ability of epithelial cancer cells to activate normal fibroblasts may provide a new anticancer therapeutic strategy.
Keywords: cancer-associated fibroblasts; collagen remodeling; colorectal cancer cells; epithelial/mesenchymal state; fibroblast activation.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures






Similar articles
-
Exosomes Derived from the Human Primary Colorectal Cancer Cell Line SW480 Orchestrate Fibroblast-Led Cancer Invasion.Proteomics. 2020 Jul;20(14):e2000016. doi: 10.1002/pmic.202000016. Proteomics. 2020. PMID: 32438511
-
Fluid Shear Stress Induces EMT of Circulating Tumor Cells via JNK Signaling in Favor of Their Survival during Hematogenous Dissemination.Int J Mol Sci. 2020 Oct 30;21(21):8115. doi: 10.3390/ijms21218115. Int J Mol Sci. 2020. PMID: 33143160 Free PMC article.
-
The impact of stromal Hic-5 on the tumorigenesis of colorectal cancer through lysyl oxidase induction and stromal remodeling.Oncogene. 2018 Mar;37(9):1205-1219. doi: 10.1038/s41388-017-0033-y. Epub 2017 Dec 15. Oncogene. 2018. PMID: 29242607
-
Expression and Function of a Disintegrin and Metalloproteinases in Cancer-Associated Fibroblasts of Colorectal Cancer.Digestion. 2020;101(1):18-24. doi: 10.1159/000504087. Epub 2019 Nov 13. Digestion. 2020. PMID: 31722362 Review.
-
Extracellular Matrix Alterations in Metastatic Processes.Int J Mol Sci. 2019 Oct 7;20(19):4947. doi: 10.3390/ijms20194947. Int J Mol Sci. 2019. PMID: 31591367 Free PMC article. Review.
Cited by
-
Biophysical Parameters Can Induce Epithelial-to-Mesenchymal Phenotypic and Genotypic Changes in HT-29 Cells: A Preliminary Study.Int J Mol Sci. 2023 Feb 16;24(4):3956. doi: 10.3390/ijms24043956. Int J Mol Sci. 2023. PMID: 36835368 Free PMC article.
-
Distinct evolutionary patterns of tumour-immune escape and elimination determined by extracellular matrix architectures.J R Soc Interface. 2025 Jul;22(228):20250116. doi: 10.1098/rsif.2025.0116. Epub 2025 Jul 9. J R Soc Interface. 2025. PMID: 40631902 Free PMC article.
-
Studying breast cancer lung metastasis using a multi-compartment microfluidic device with a mimetic tumor-stroma interaction model.Transl Oncol. 2025 Mar;53:102303. doi: 10.1016/j.tranon.2025.102303. Epub 2025 Feb 4. Transl Oncol. 2025. PMID: 39904278 Free PMC article.
-
The mechanobiology of fibroblast activation in disease.APL Bioeng. 2025 Jun 18;9(2):021505. doi: 10.1063/5.0272393. eCollection 2025 Jun. APL Bioeng. 2025. PMID: 40538750 Free PMC article. Review.
-
Monitoring the Intracellular pH and Metabolic State of Cancer Cells in Response to Chemotherapy Using a Combination of Phosphorescence Lifetime Imaging Microscopy and Fluorescence Lifetime Imaging Microscopy.Int J Mol Sci. 2023 Dec 19;25(1):49. doi: 10.3390/ijms25010049. Int J Mol Sci. 2023. PMID: 38203221 Free PMC article.
References
-
- Chopra V., Sangarappillai R.M., Romero-Canelón I., Jones A.M. Lysyl Oxidase Like-2 (LOXL2): An Emerging Oncology Target. Adv. Ther. 2020;3:1900119. doi: 10.1002/adtp.201900119. - DOI
-
- Sahai E., Astsaturov I., Cukierman E., DeNardo D.G., Egeblad M., Evans R.M., Fearon D., Greten F.R., Hingorani S.R., Hunter T., et al. A framework for advancing our understanding of cancer-associated fibroblasts. Nat. Rev. Cancer. 2020;20:174–186. doi: 10.1038/s41568-019-0238-1. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
