

GTP-State-Selective Cyclic Peptide Ligands of K-Ras(G12D) Block Its Interaction with Raf
- PMID: 33145412
- PMCID: PMC7596874
- DOI: 10.1021/acscentsci.0c00514
GTP-State-Selective Cyclic Peptide Ligands of K-Ras(G12D) Block Its Interaction with Raf
Abstract
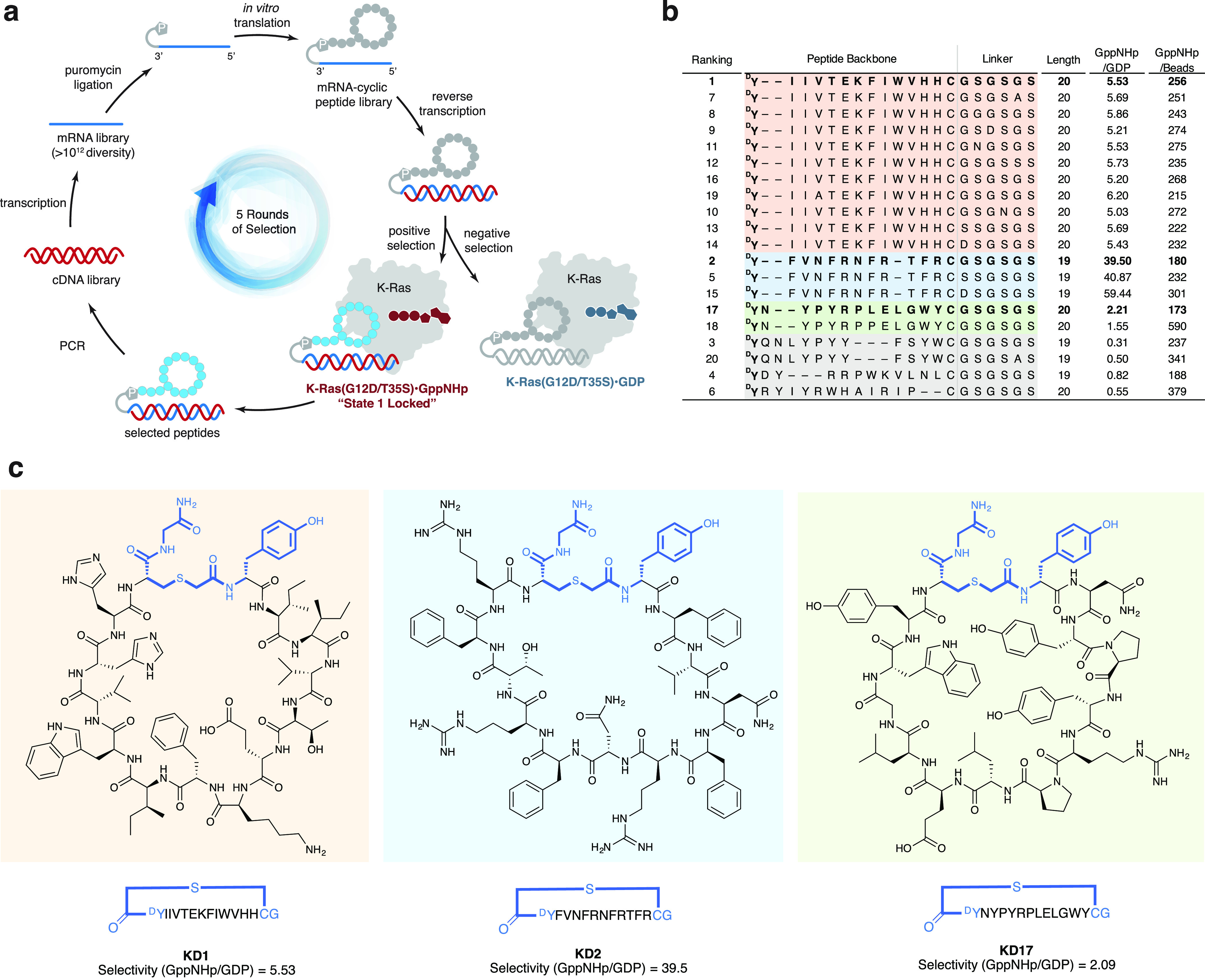
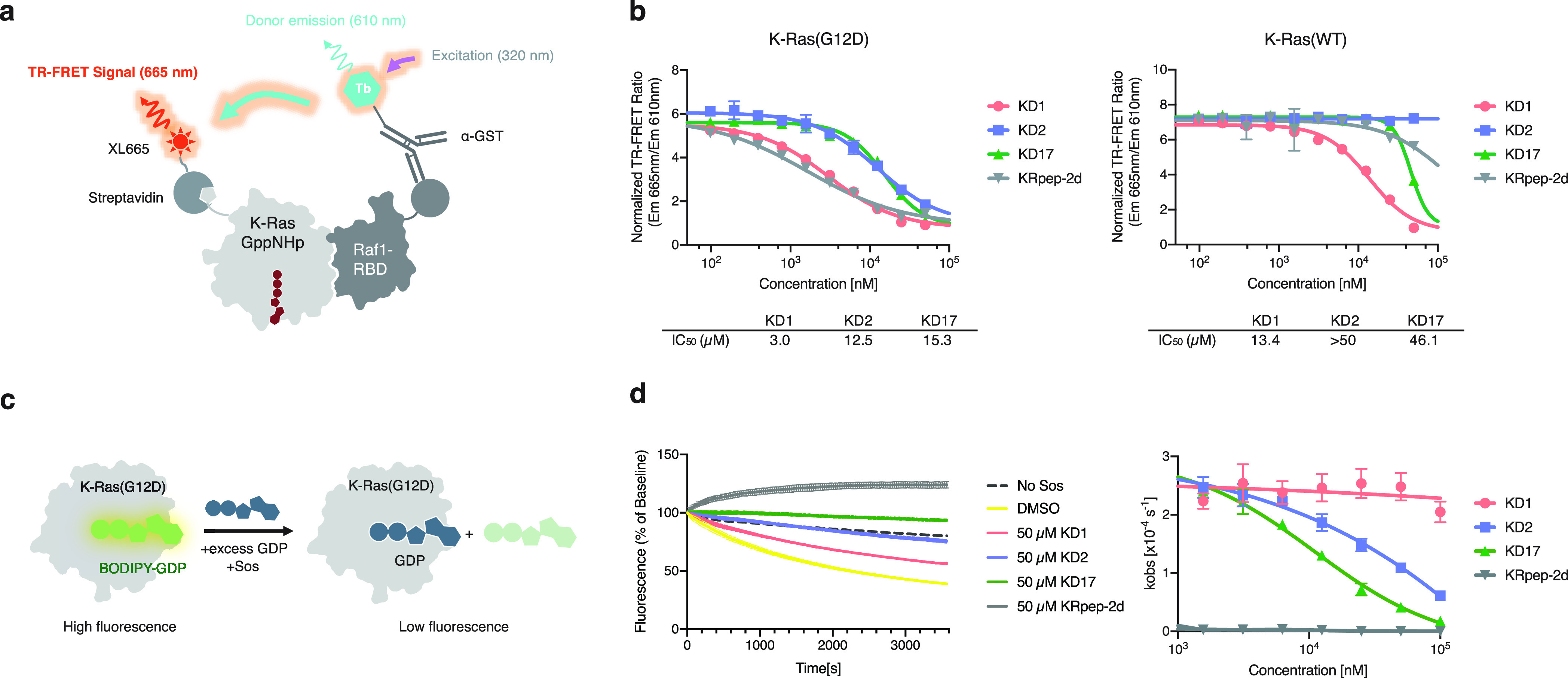
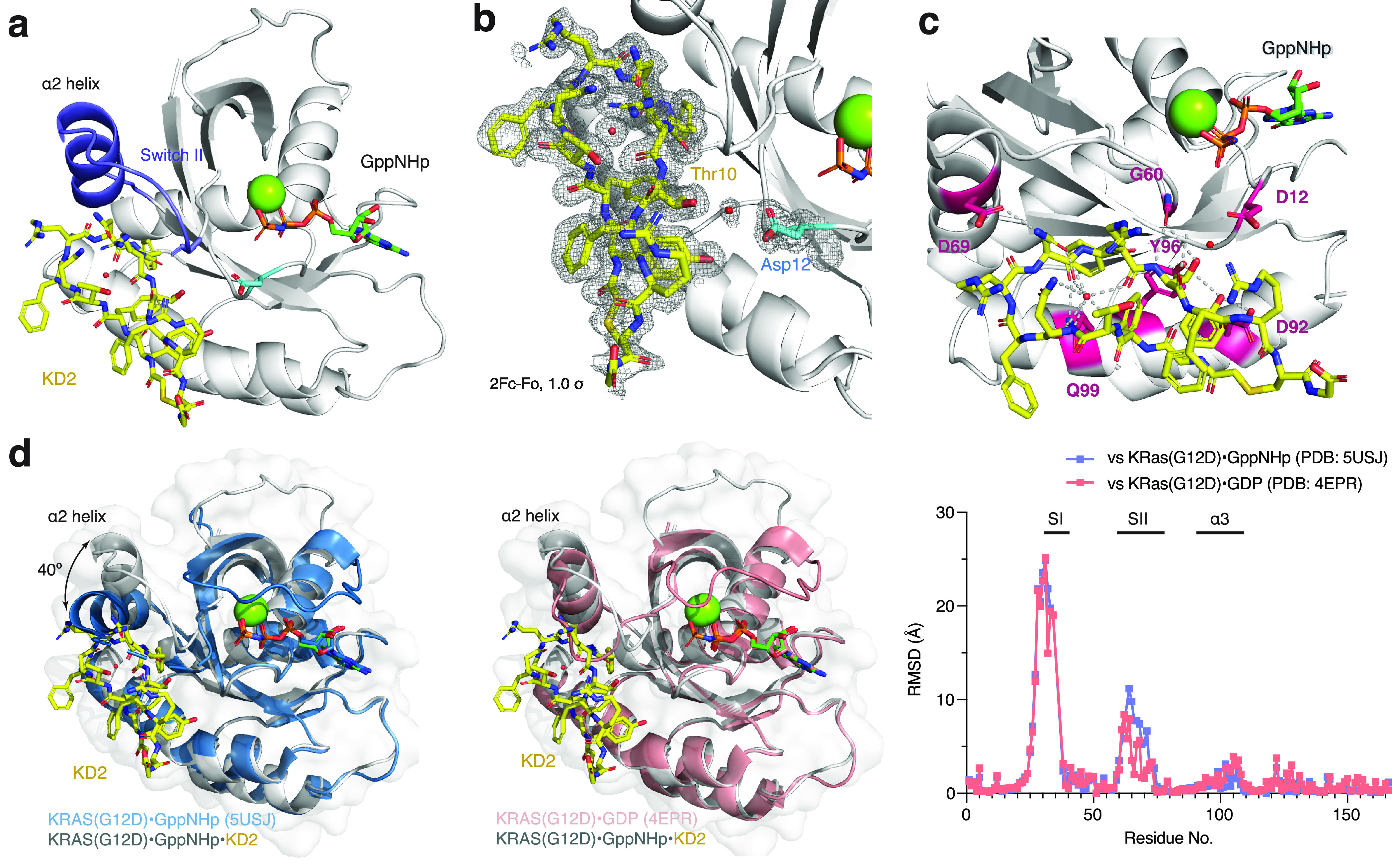
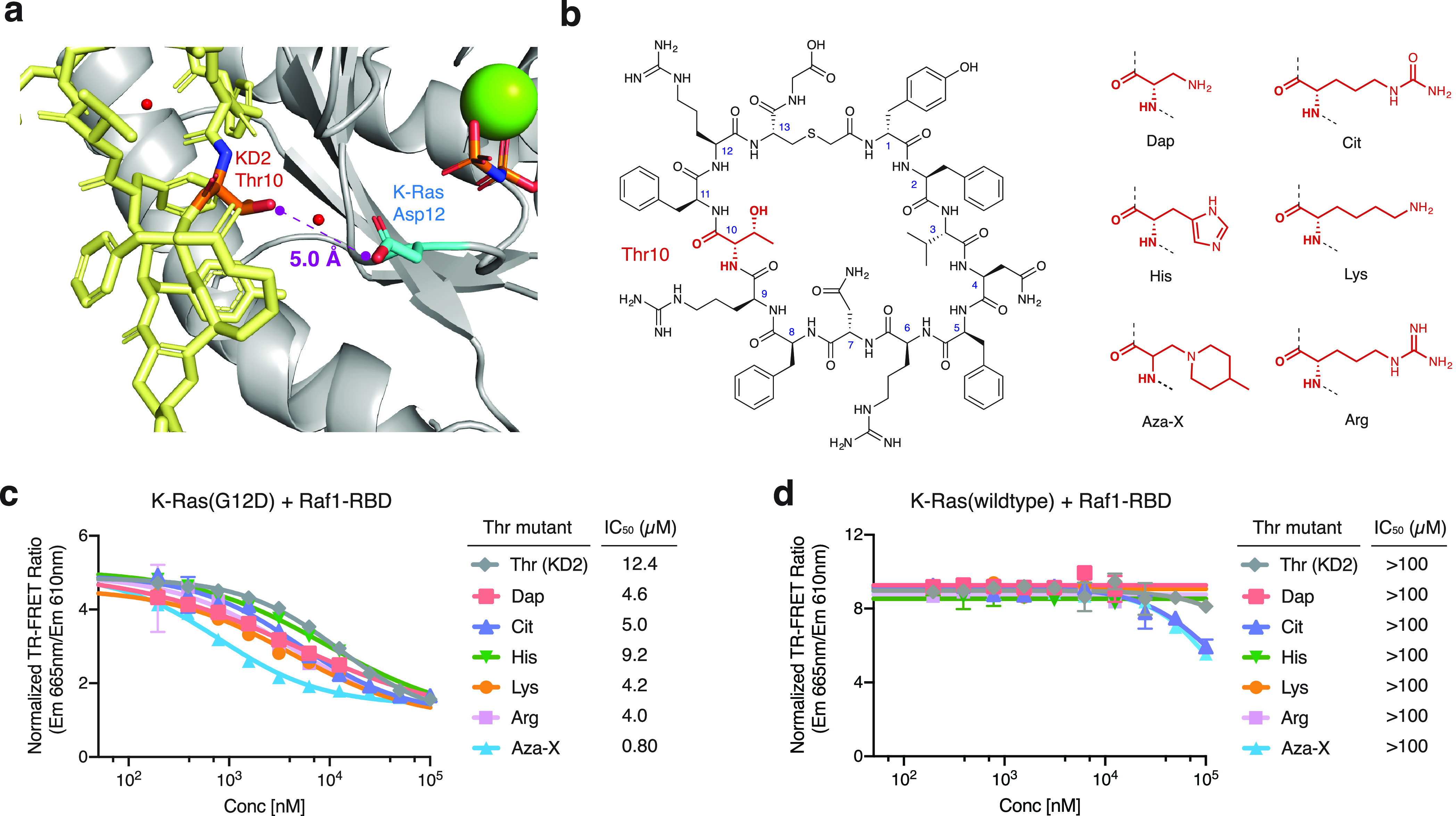
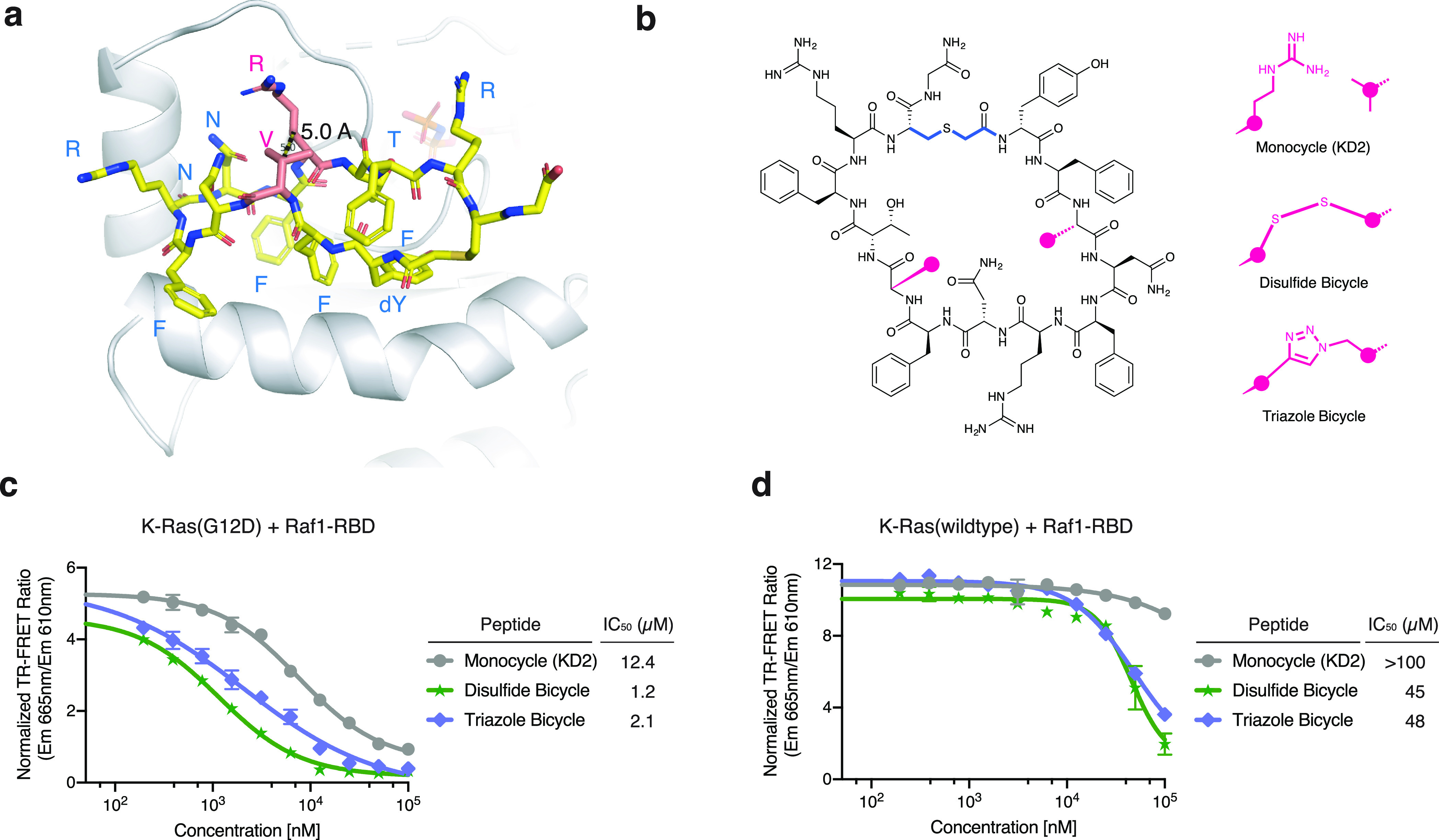
We report the identification of three cyclic peptide ligands of K-Ras(G12D) using an integrated in vitro translation-mRNA display selection platform. These cyclic peptides show preferential binding to the GTP-bound state of K-Ras(G12D) over the GDP-bound state and block Ras-Raf interaction. A co-crystal structure of peptide KD2 with K-Ras(G12D)·GppNHp reveals that this peptide binds in the Switch II groove region with concomitant opening of the Switch II loop and a 40° rotation of the α2 helix, and that a threonine residue (Thr10) on KD2 has direct access to the mutant aspartate (Asp12) on K-Ras. Replacing this threonine with non-natural amino acids afforded peptides with improved potency at inhibiting the interaction between Raf1-RBD and K-Ras(G12D) but not wildtype K-Ras. The union of G12D over wildtype selectivity and GTP state/GDP state selectivity is particularly desirable, considering that oncogenic K-Ras(G12D) exists predominantly in the GTP state in cancer cells, and wildtype K-Ras signaling is important for the maintenance of healthy cells.
Conflict of interest statement
The authors declare the following competing financial interest(s): Kevan Shokat has consulting agreements for the following companies involving cash and/or stock compensation: Black Diamond Therapeutics, BridGene Biosciences, Denali Therapeutics, Dice Molecules, eFFECTOR Therapeutics, Erasca, Genentech/Roche, Janssen Pharmaceuticals, Kumquat Biosciences, Kura Oncology, Merck, Mitokinin, Petra Pharma, Qulab Inc. Revolution Medicines, Type6 Therapeutics, Venthera, Wellspring Biosciences (Araxes Pharma).
Figures
Similar articles
-
Crystal Structure of a Human K-Ras G12D Mutant in Complex with GDP and the Cyclic Inhibitory Peptide KRpep-2d.ACS Med Chem Lett. 2017 May 10;8(7):732-736. doi: 10.1021/acsmedchemlett.7b00128. eCollection 2017 Jul 13. ACS Med Chem Lett. 2017. PMID: 28740607 Free PMC article.
-
1H, 15N backbone assignment and comparative analysis of the wild type and G12C, G12D, G12V mutants of K-Ras bound to GDP at physiological pH.Biomol NMR Assign. 2020 Apr;14(1):1-7. doi: 10.1007/s12104-019-09909-7. Epub 2019 Aug 29. Biomol NMR Assign. 2020. PMID: 31468366 Free PMC article.
-
Conformational Dynamics Allows Sampling of an "Active-like" State by Oncogenic K-Ras-GDP.J Mol Biol. 2022 Sep 15;434(17):167695. doi: 10.1016/j.jmb.2022.167695. Epub 2022 Jun 23. J Mol Biol. 2022. PMID: 35752212
-
K-Ras(G12D)-selective inhibitory peptides generated by random peptide T7 phage display technology.Biochem Biophys Res Commun. 2017 Mar 11;484(3):605-611. doi: 10.1016/j.bbrc.2017.01.147. Epub 2017 Jan 30. Biochem Biophys Res Commun. 2017. PMID: 28153726
-
Peptides That Block RAS-p21 Protein-Induced Cell Transformation.Biomedicines. 2023 Feb 6;11(2):471. doi: 10.3390/biomedicines11020471. Biomedicines. 2023. PMID: 36831007 Free PMC article. Review.
Cited by
-
Accelerated Identification of Cell Active KRAS Inhibitory Macrocyclic Peptides using Mixture Libraries and Automated Ligand Identification System (ALIS) Technology.J Med Chem. 2022 Jul 14;65(13):8961-8974. doi: 10.1021/acs.jmedchem.2c00154. Epub 2022 Jun 15. J Med Chem. 2022. PMID: 35707970 Free PMC article.
-
Small-Molecule Inhibitors and Degraders Targeting KRAS-Driven Cancers.Int J Mol Sci. 2021 Nov 9;22(22):12142. doi: 10.3390/ijms222212142. Int J Mol Sci. 2021. PMID: 34830024 Free PMC article. Review.
-
Oncogenic KRAS-Induced Feedback Inflammatory Signaling in Pancreatic Cancer: An Overview and New Therapeutic Opportunities.Cancers (Basel). 2021 Oct 31;13(21):5481. doi: 10.3390/cancers13215481. Cancers (Basel). 2021. PMID: 34771644 Free PMC article. Review.
-
Selected Approaches to Disrupting Protein-Protein Interactions within the MAPK/RAS Pathway.Int J Mol Sci. 2023 Apr 17;24(8):7373. doi: 10.3390/ijms24087373. Int J Mol Sci. 2023. PMID: 37108538 Free PMC article. Review.
-
Motif-guided identification of KRAS-interacting proteins.BMC Biol. 2024 Nov 19;22(1):264. doi: 10.1186/s12915-024-02067-w. BMC Biol. 2024. PMID: 39563372 Free PMC article.
References
-
- Taveras A. G.; Remiszewski S. W.; Doll R. J.; Cesarz D.; Huang E. C.; Kirschmeier P.; Pramanik B. N.; Snow M. E.; Wang Y. S.; Del Rosario J. D.; Vibulbhan B.; Bauer B. B.; Brown J. E.; Carr D.; Catino J.; Evans C. A.; Girijavallabhan V.; Heimark L.; James L.; Liberles S.; Nash C.; Perkins L.; Senior M. M.; Tsarbopoulos A.; Ganguly A. K.; Aust R.; Brown E.; Delisle D.; Fuhrman S.; Hendrickson T.; Kissinger C.; Love R.; Sisson W.; Villafranca E.; Webber S. E. Ras Oncoprotein Inhibitors: The Discovery of Potent, Ras Nucleotide Exchange Inhibitors and the Structural Determination of a Drug-Protein Complex. Bioorg. Med. Chem. 1997, 5 (1), 125–133. 10.1016/S0968-0896(96)00202-7. - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
