

Fully human anti-CD39 antibody potently inhibits ATPase activity in cancer cells via uncompetitive allosteric mechanism
- PMID: 33146056
- PMCID: PMC7646477
- DOI: 10.1080/19420862.2020.1838036
Fully human anti-CD39 antibody potently inhibits ATPase activity in cancer cells via uncompetitive allosteric mechanism
Abstract
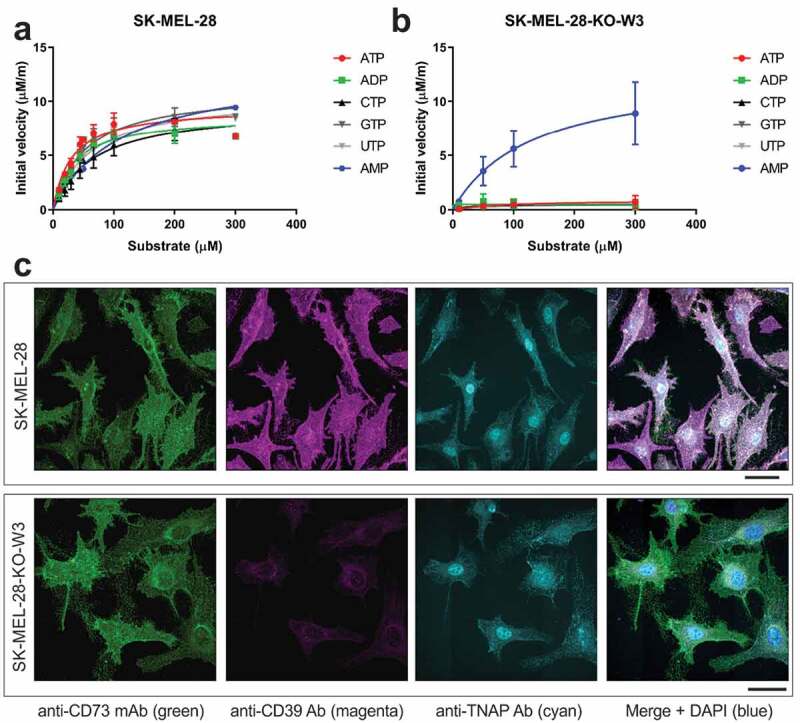
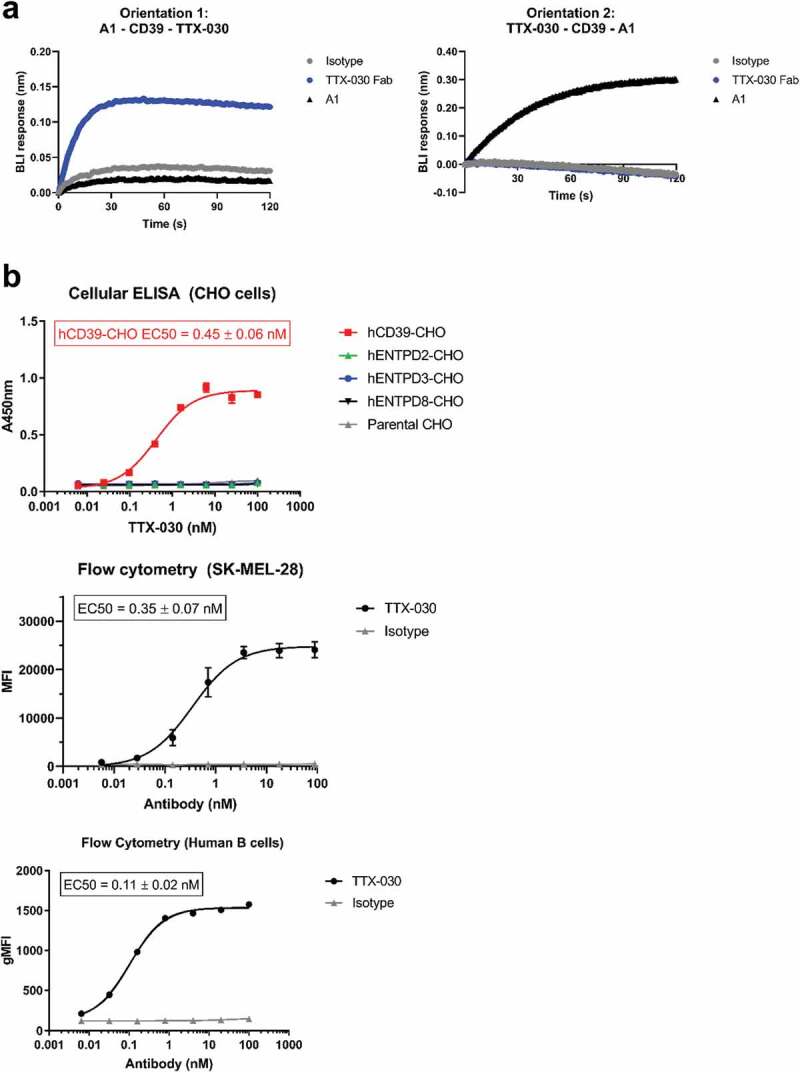
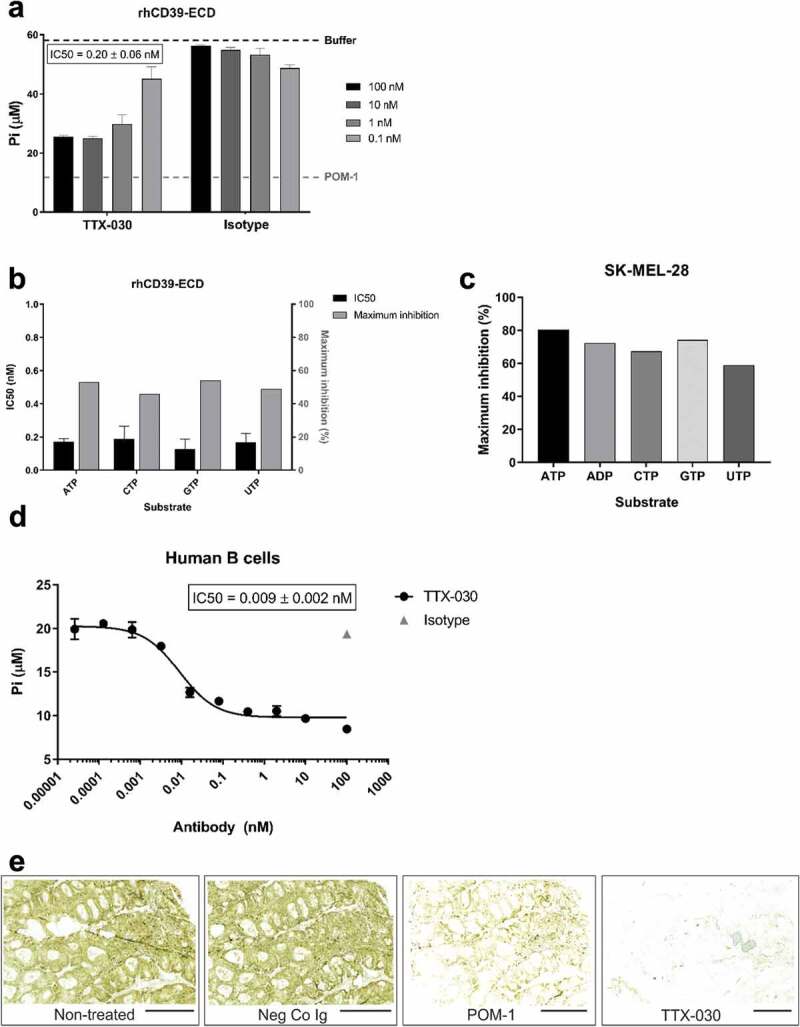
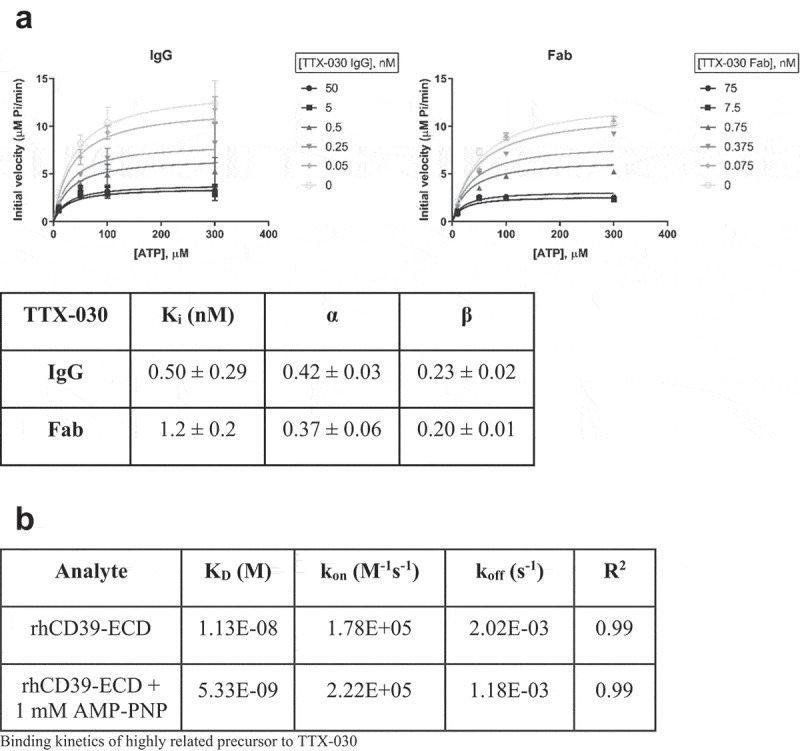
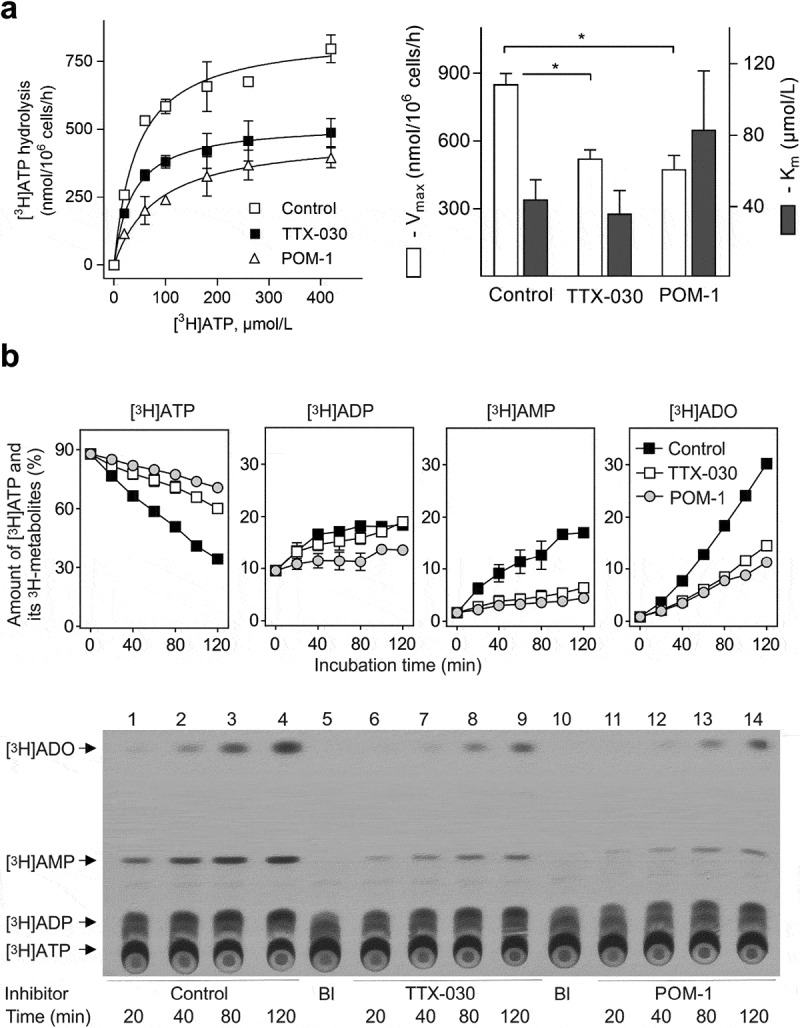
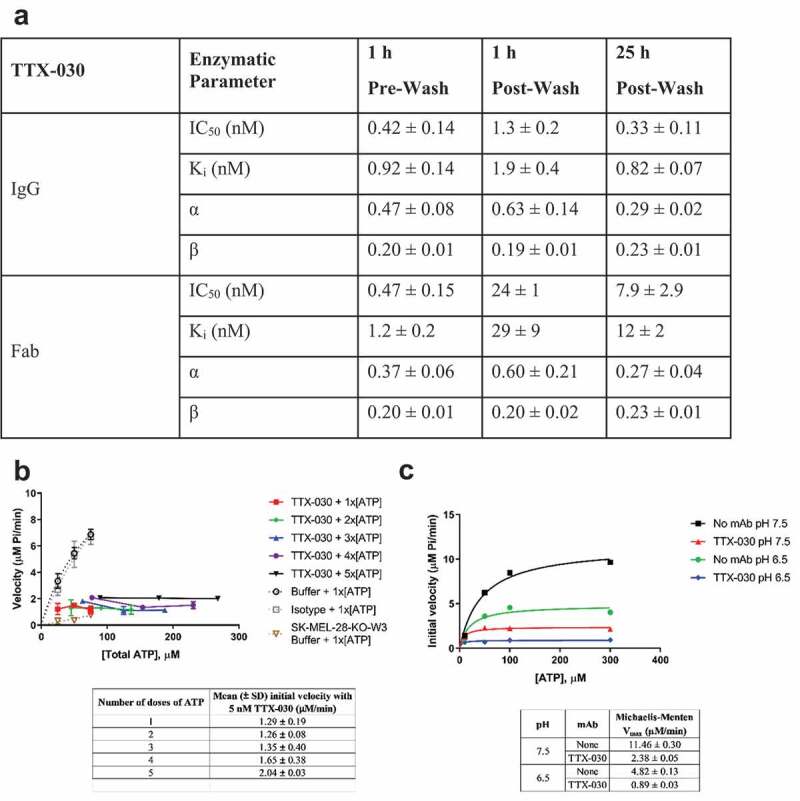
The extracellular ATP/adenosine axis in the tumor microenvironment (TME) has emerged as an important immune-regulatory pathway. Nucleoside triphosphate diphosphohydrolase-1 (NTPDase1), otherwise known as CD39, is highly expressed in the TME, both on infiltrating immune cells and tumor cells across a broad set of cancer indications. CD39 processes pro-inflammatory extracellular ATP to ADP and AMP, which is then processed by Ecto-5'-nucleotidase/CD73 to immunosuppressive adenosine. Directly inhibiting the enzymatic function of CD39 via an antibody has the potential to unleash an immune-mediated anti-tumor response via two mechanisms: 1) increasing the availability of immunostimulatory extracellular ATP released by damaged and/or dying cells, and 2) reducing the generation and accumulation of suppressive adenosine within the TME. Tizona Therapeutics has engineered a novel first-in-class fully human anti-CD39 antibody, TTX-030, that directly inhibits CD39 ATPase enzymatic function with sub-nanomolar potency. Further characterization of the mechanism of inhibition by TTX-030 using CD39+ human melanoma cell line SK-MEL-28 revealed an uncompetitive allosteric mechanism (α < 1). The uncompetitive mechanism of action enables TTX-030 to inhibit CD39 at the elevated ATP concentrations reported in the TME. Maximal inhibition of cellular CD39 ATPase velocity was 85%, which compares favorably to results reported for antibody inhibitors to other enzyme targets. The allosteric mechanism of TTX-030 was confirmed via mapping the epitope to a region of CD39 distant from its active site, which suggests possible models for how potent inhibition is achieved. In summary, TTX-030 is a potent allosteric inhibitor of CD39 ATPase activity that is currently being evaluated in clinical trials for cancer therapy.
Keywords: ATP; CD39; allosteric regulation; cancer; cancer therapy; enzyme inhibitor; enzyme kinetics; enzyme mechanism; monoclonal antibody.
Conflict of interest statement
Spatola, BN, Lerner, AG, Wong, C, dela Cruz, T, Welch, M, Fung, W, Kovalenko, M, Beers, C, Corbin, J, and Soros, VB are, or were, employees of Tizona Therapeutics and hold stock and/or stock options in the same. Research by Losenkova, K and Yegutkin, GG was in part sponsored by Tizona Therapeutics.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials