

Role of Aberrant Spontaneous Neurotransmission in SNAP25-Associated Encephalopathies
- PMID: 33147442
- PMCID: PMC7790958
- DOI: 10.1016/j.neuron.2020.10.012
Role of Aberrant Spontaneous Neurotransmission in SNAP25-Associated Encephalopathies
Abstract
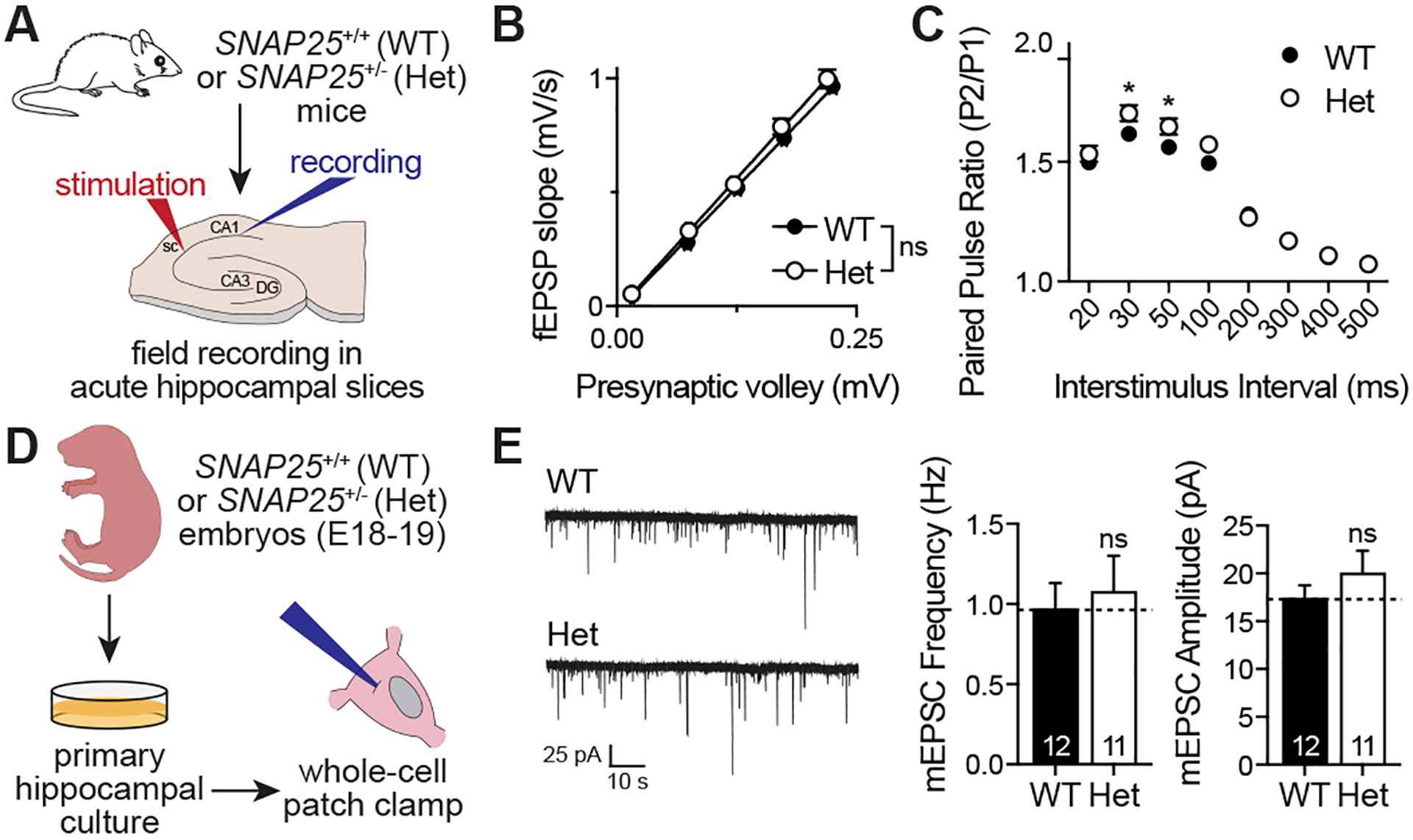
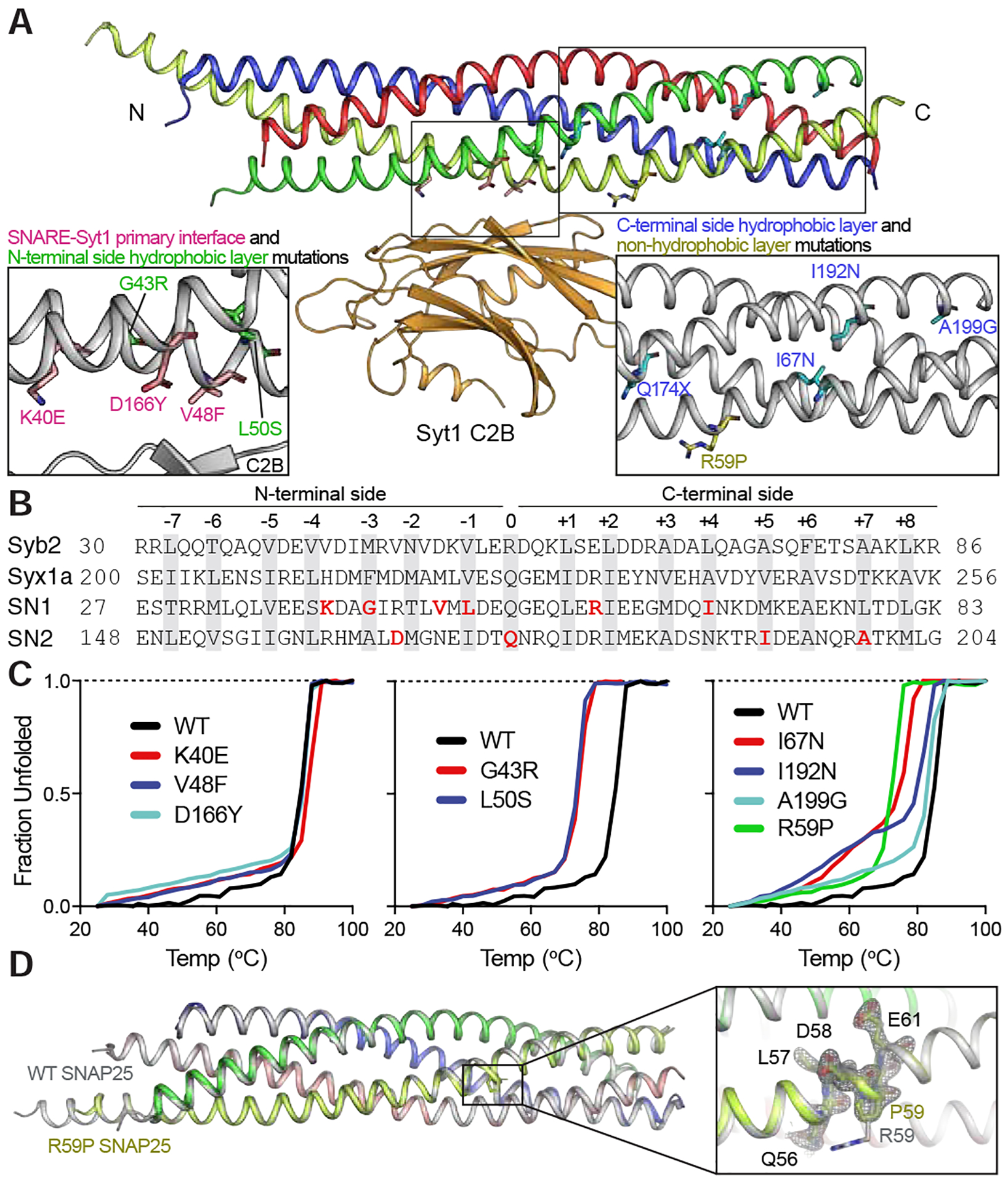
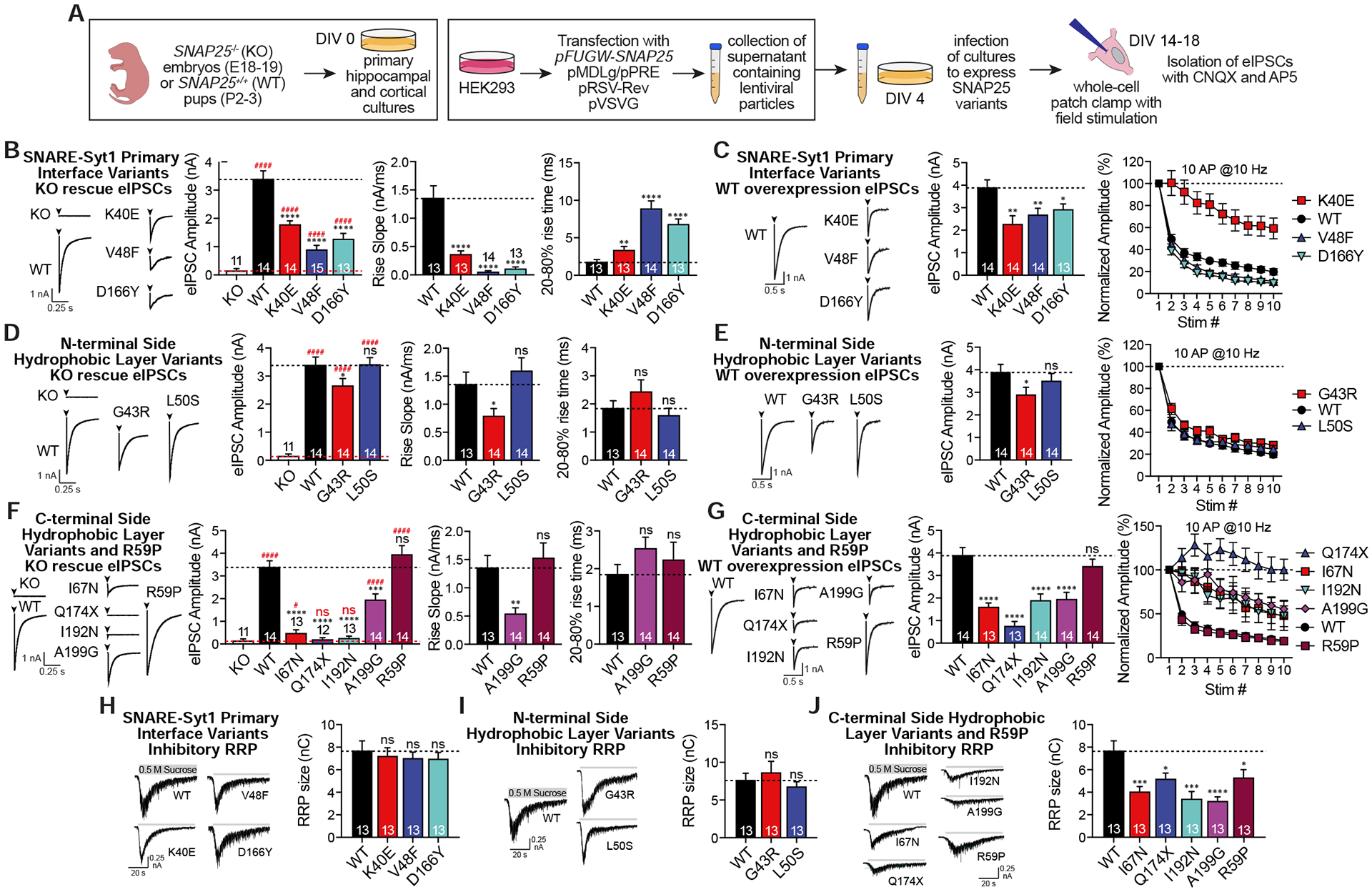
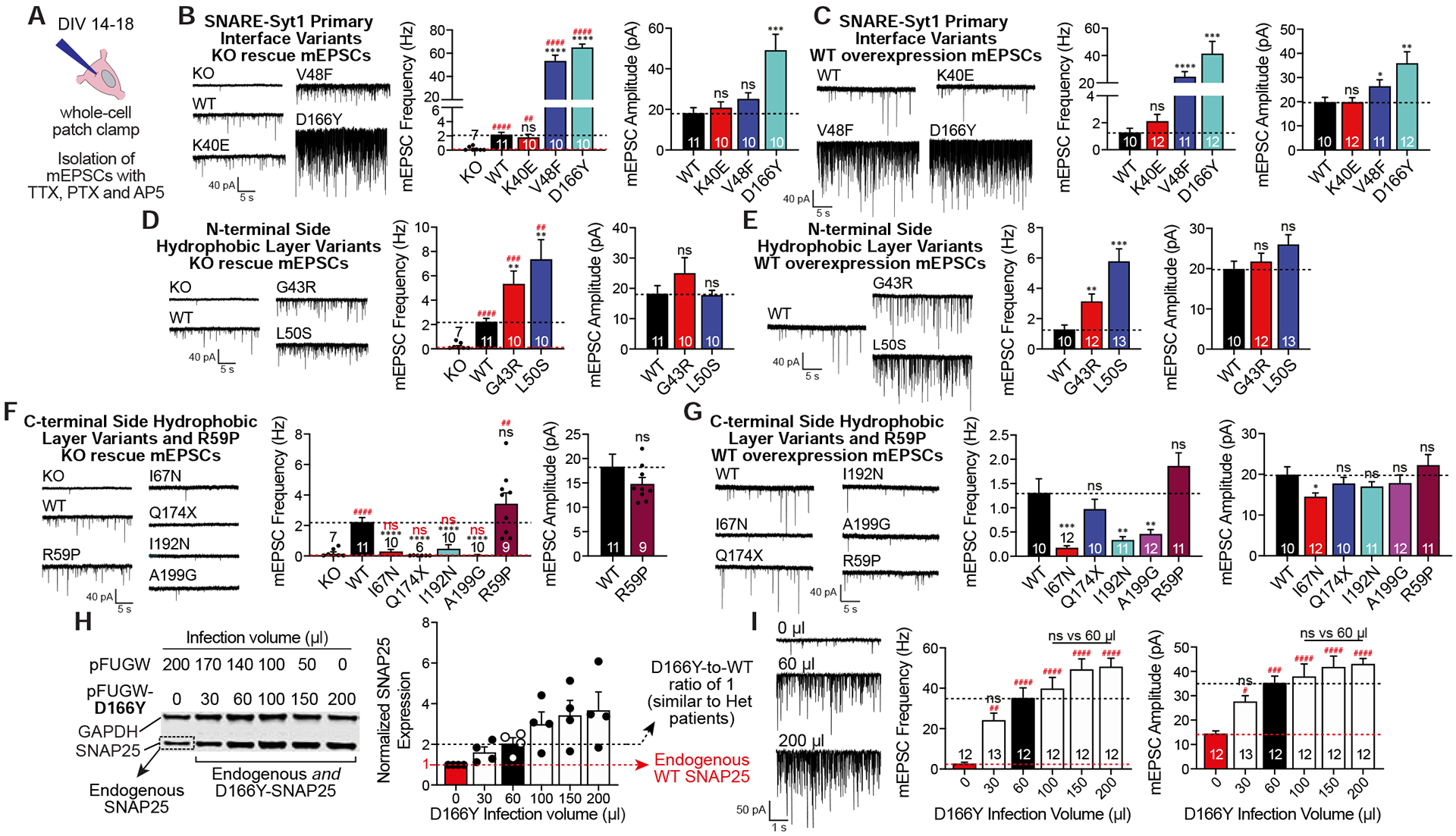
SNARE (soluble N-ethylmaleimide sensitive factor attachment protein receptor) complex, composed of synaptobrevin, syntaxin, and SNAP25, forms the essential fusion machinery for neurotransmitter release. Recent studies have reported several mutations in the gene encoding SNAP25 as a causative factor for developmental and epileptic encephalopathies of infancy and childhood with diverse clinical manifestations. However, it remains unclear how SNAP25 mutations give rise to these disorders. Here, we show that although structurally clustered mutations in SNAP25 give rise to related synaptic transmission phenotypes, specific alterations in spontaneous neurotransmitter release are a key factor to account for disease heterogeneity. Importantly, we identified a single mutation that augments spontaneous release without altering evoked release, suggesting that aberrant spontaneous release is sufficient to cause disease in humans.
Keywords: SNAP25; child neurology; development; developmental delay; epilepsy; exocytosis; neurotransmitter; spontaneous release; synaptic transmission; synaptotagmin.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
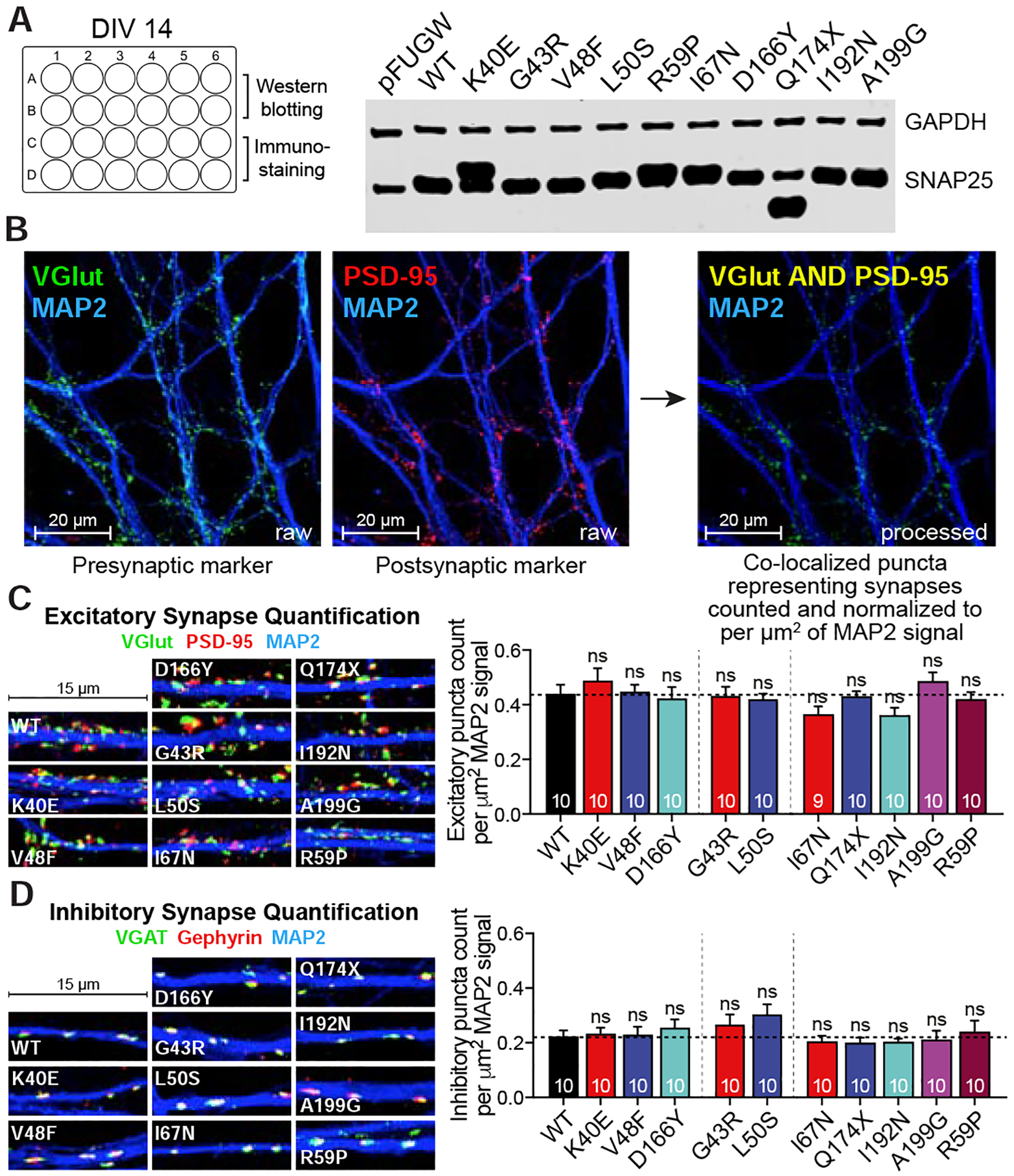
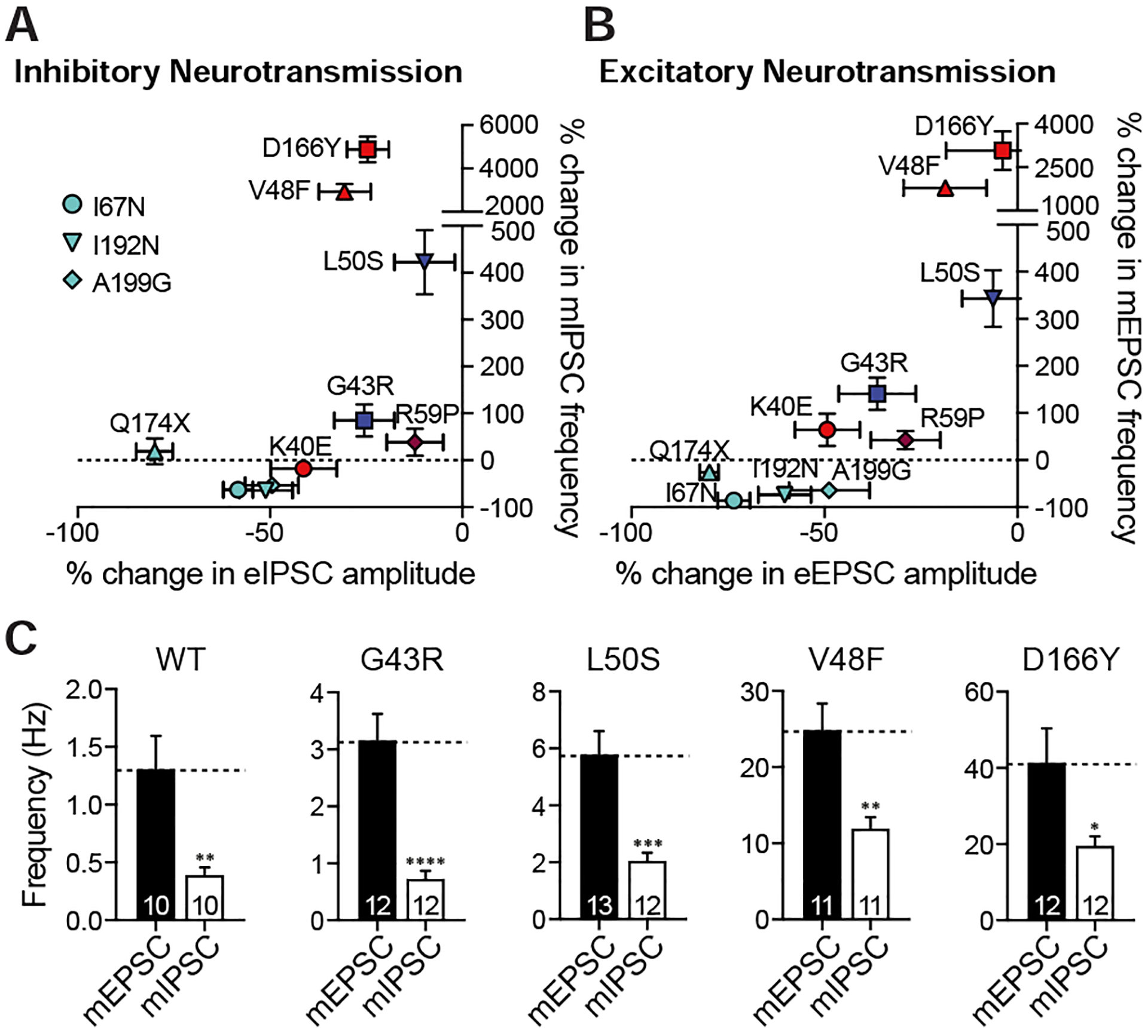
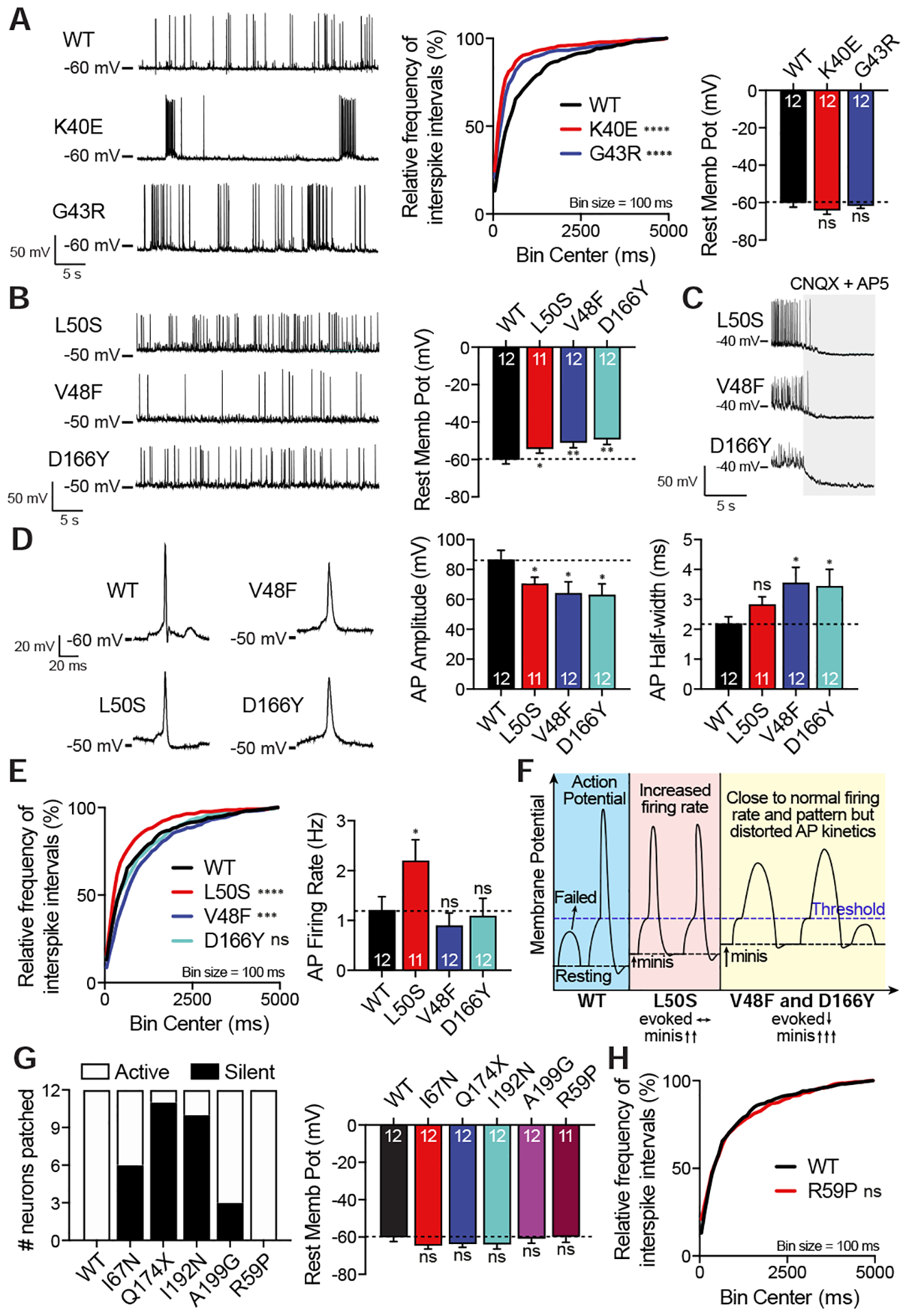
Comment in
-
Molecule-to-Circuit Disease Mechanisms of a Synaptic SNAREopathy.Neuron. 2021 Jan 6;109(1):1-3. doi: 10.1016/j.neuron.2020.12.009. Neuron. 2021. PMID: 33412092
References
-
- Adams PD, Grosse-Kunstleve RW, Hung LW, Ioerger TR, McCoy AJ, Moriarty NW, Read RJ, Sacchettini JC, Sauter NK, and Terwilliger TC (2002). PHENIX: building new software for automated crystallographic structure determination. Acta Crystallogr D Biol Crystallogr 58, 1948–1954. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
