

GDF6-CD99 Signaling Regulates Src and Ewing Sarcoma Growth
- PMID: 33147457
- PMCID: PMC7688343
- DOI: 10.1016/j.celrep.2020.108332
GDF6-CD99 Signaling Regulates Src and Ewing Sarcoma Growth
Abstract
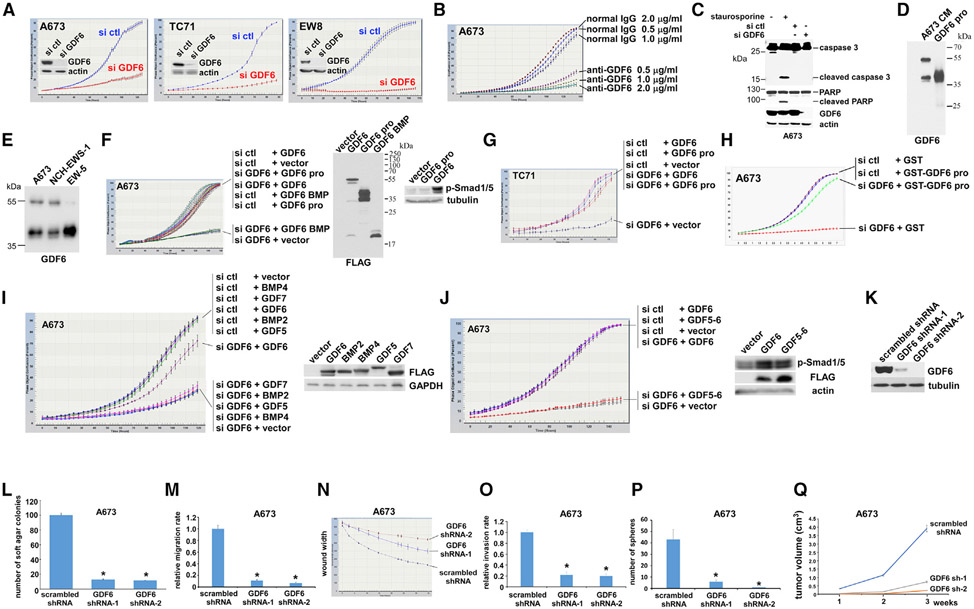
We report here that the autocrine signaling mediated by growth and differentiation factor 6 (GDF6), a member of the bone morphogenetic protein (BMP) family of cytokines, maintains Ewing sarcoma growth by preventing Src hyperactivation. Surprisingly, Ewing sarcoma depends on the prodomain, not the BMP domain, of GDF6. We demonstrate that the GDF6 prodomain is a ligand for CD99, a transmembrane protein that has been widely used as a marker of Ewing sarcoma. The binding of the GDF6 prodomain to the CD99 extracellular domain results in recruitment of CSK (C-terminal Src kinase) to the YQKKK motif in the intracellular domain of CD99, inhibiting Src activity. GDF6 silencing causes hyperactivation of Src and p21-dependent growth arrest. We demonstrate that two GDF6 prodomain mutants linked to Klippel-Feil syndrome are hyperactive in CD99-Src signaling. These results reveal a cytokine signaling pathway that regulates the CSK-Src axis and cancer cell proliferation and suggest the gain-of-function activity for disease-causing GDF6 mutants.
Keywords: CD99; CSK; Ewing sarcoma; GDF6; Klippel-Feil syndrome; Src; proteomics; secretome.
Copyright © 2020 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures




References
-
- Ambros IM, Ambros PF, Strehl S, Kovar H, Gadner H, and Salzer-Kuntschik M (1991). MIC2 is a specific marker for Ewing’s sarcoma and peripheral primitive neuroectodermal tumors. Evidence for a common histogenesis of Ewing’s sarcoma and peripheral primitive neuroectodermal tumors from MIC2 expression and specific chromosome aberration. Cancer 67, 1886–1893. - PubMed
-
- Aono A, Hazama M, Notoya K, Taketomi S, Yamasaki H, Tsukuda R, Sasaki S, and Fujisawa Y (1995). Potent ectopic bone-inducing activity of bone morphogenetic protein-4/7 heterodimer. Biochem. Biophys. Res. Commun 210, 670–677. - PubMed
-
- Arbet-Engels C, Tartare-Deckert S, and Eckhart W (1999). C-terminal Src kinase associates with ligand-stimulated insulin-like growth factor-I receptor. J. Biol. Chem 274, 5422–5428. - PubMed
-
- Asai-Coakwell M, March L, Dai XH, Duval M, Lopez I, French CR, Famulski J, De Baere E, Francis PJ, Sundaresan P, et al. (2013). Contribution of growth differentiation factor 6-dependent cell survival to early-onset retinal dystrophies. Hum. Mol. Genet 22, 1432–1442. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
