

Lipid Droplets in Brown Adipose Tissue Are Dispensable for Cold-Induced Thermogenesis
- PMID: 33147469
- PMCID: PMC7696656
- DOI: 10.1016/j.celrep.2020.108348
Lipid Droplets in Brown Adipose Tissue Are Dispensable for Cold-Induced Thermogenesis
Abstract
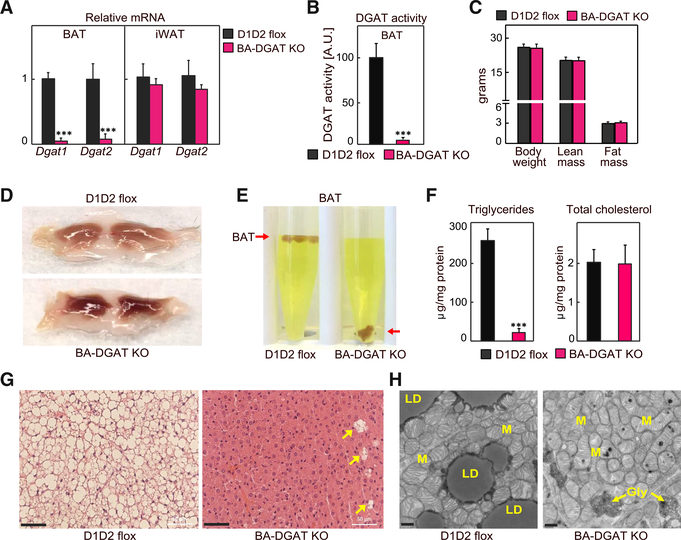
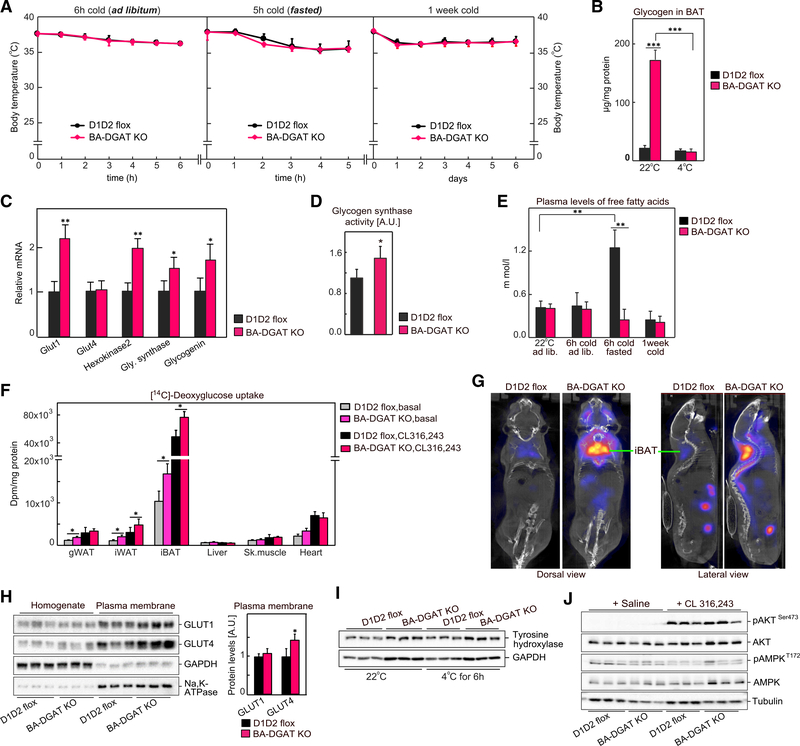
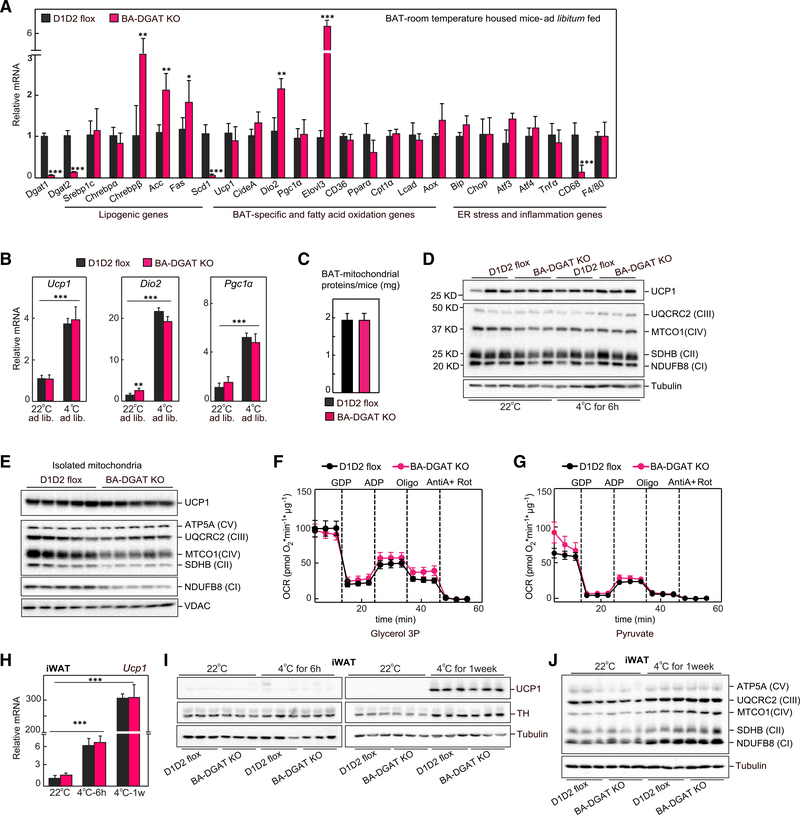
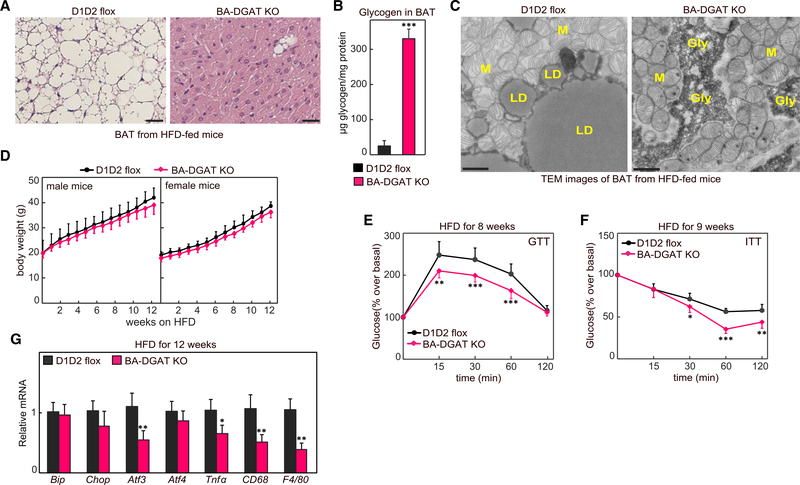
Brown adipocytes store metabolic energy as triglycerides (TGs) in lipid droplets (LDs). Fatty acids released from brown adipocyte LDs by lipolysis are thought to activate and fuel UCP1-mediated thermogenesis. Here, we test this hypothesis by preventing fatty acid storage in murine brown adipocytes through brown adipose tissue (BAT)-specific deletions of the TG synthesis enzymes DGAT1 and DGAT2 (BA-DGAT KO). Despite the absence of TGs in brown adipocytes, BAT is functional, and BA-DGAT-KO mice maintain euthermia during acute or chronic cold exposure. As apparent adaptations to the lack of TG, brown adipocytes of BA-DGAT-KO mice appear to use circulating glucose and fatty acids, and stored glycogen, to fuel thermogenesis. Moreover, BA-DGAT-KO mice are resistant to diet-induced glucose intolerance, likely because of increased glucose disposal by BAT. We conclude that TGs in BAT are dispensable for its contribution to cold-induced thermogenesis, at least when other fuel sources are available.
Keywords: Brown adipose tissue; DGAT1; DGAT2; cold-induced thermogenesis; glucose intolerance; lipid droplets; triglycerides.
Copyright © 2020 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests T.C.W. has an immediate family member working for Pfizer, Inc. T.C.W. and R.V.F. have a sponsored research agreement with Pfizer, Inc.
Figures
References
-
- Bartelt A, Bruns OT, Reimer R, Hohenberg H, Ittrich H, Peldschus K, Kaul MG, Tromsdorf UI, Weller H, Waurisch C, et al. (2011). Brown adipose tissue activity controls triglyceride clearance. Nat. Med 17, 200–205. - PubMed
-
- Bertholet AM, and Kirichok Y (2019). The mechanism FA-dependent H(+) transport by UCP1. Handb. Exp. Pharmacol 251, 143–159. - PubMed
-
- Bertholet AM, Kazak L, Chouchani ET, Bogaczynska MG, Paranjpe I, Wainwright GL, Betourne A, Kajimura S, Spiegelman BM, and Kirichok Y (2017). Mitochondrial patch clamp of beige adipocytes reveals UCP1-positive and UCP1-negative cells both exhibiting futile creatine cycling. Cell Metab. 25, 811–822.e4. - PMC - PubMed
-
- Cannon B, and Nedergaard J (2004). Brown adipose tissue: function and physiological significance. Physiol. Rev. 84, 277–359. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
