

Pyruvate Kinase Controls Signal Strength in the Insulin Secretory Pathway
- PMID: 33147484
- PMCID: PMC7685238
- DOI: 10.1016/j.cmet.2020.10.007
Pyruvate Kinase Controls Signal Strength in the Insulin Secretory Pathway
Abstract
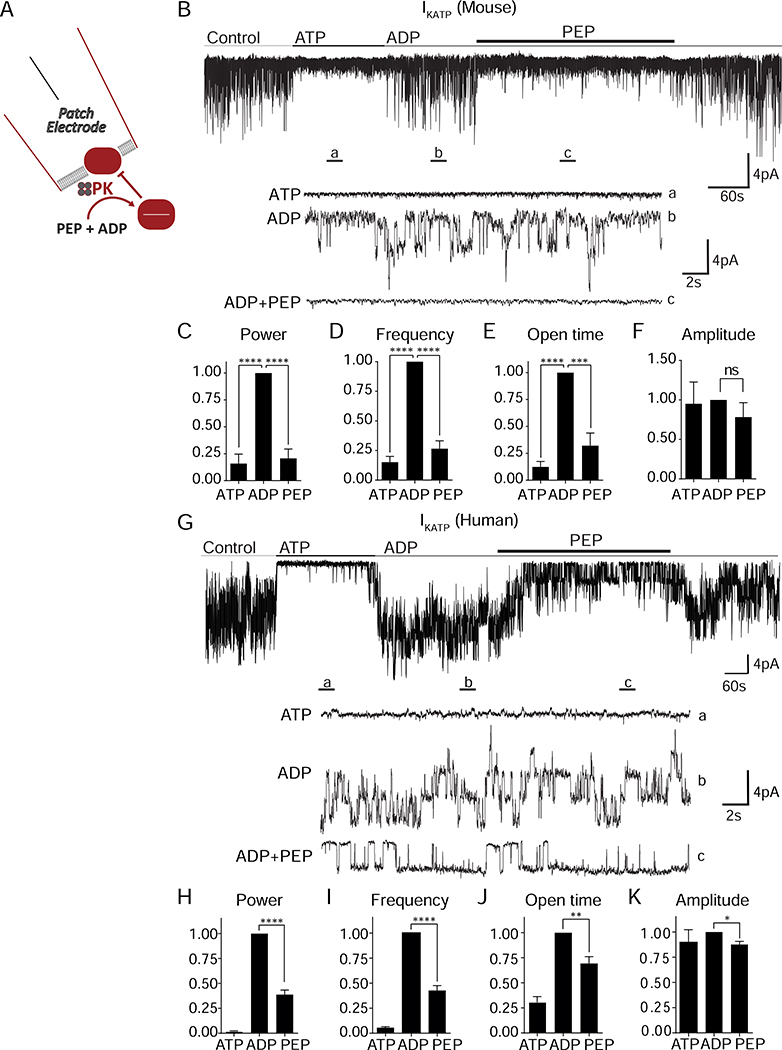
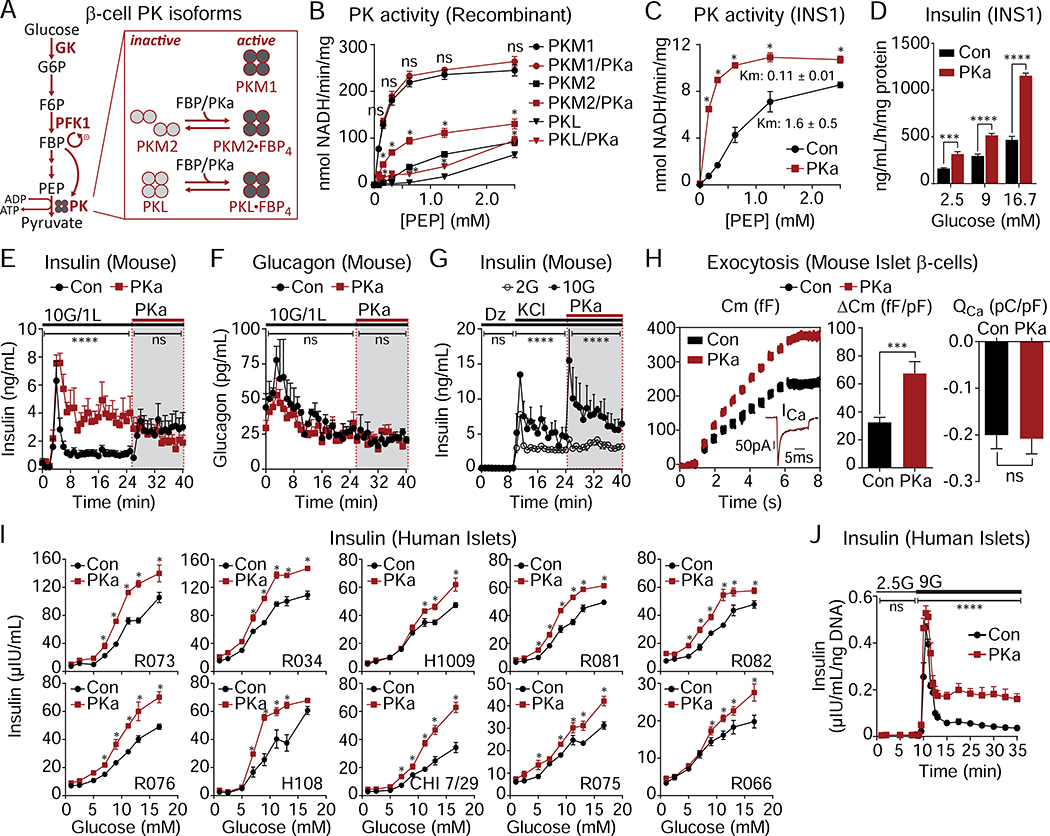
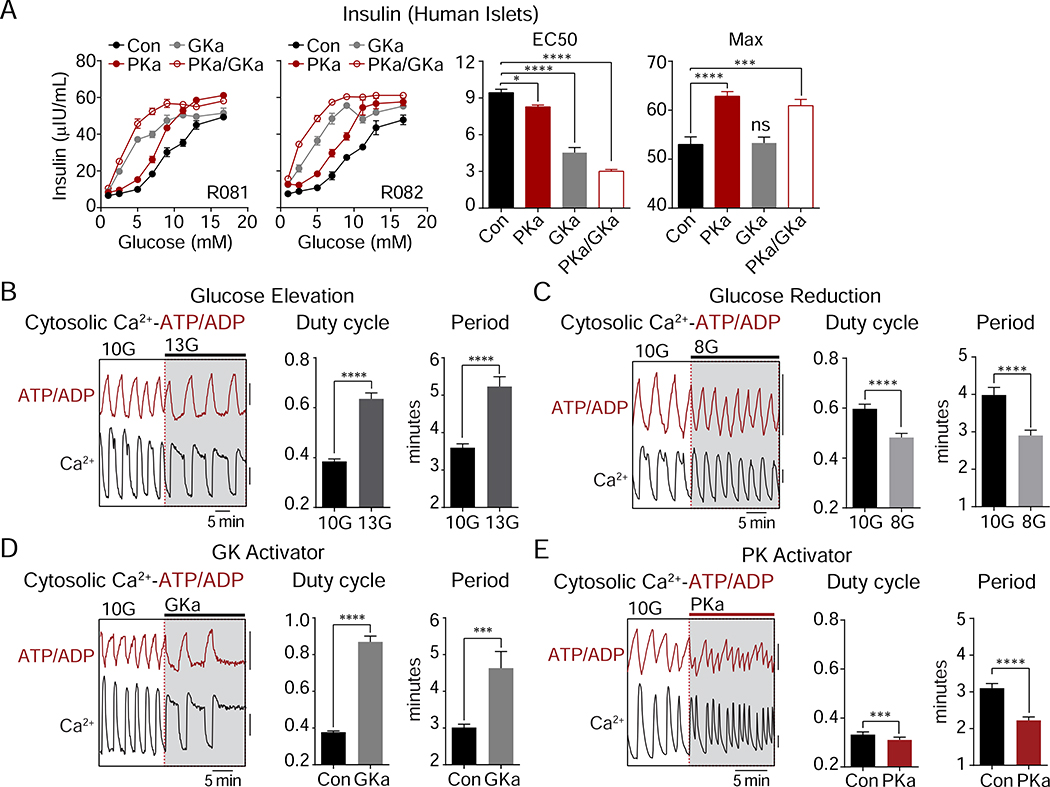
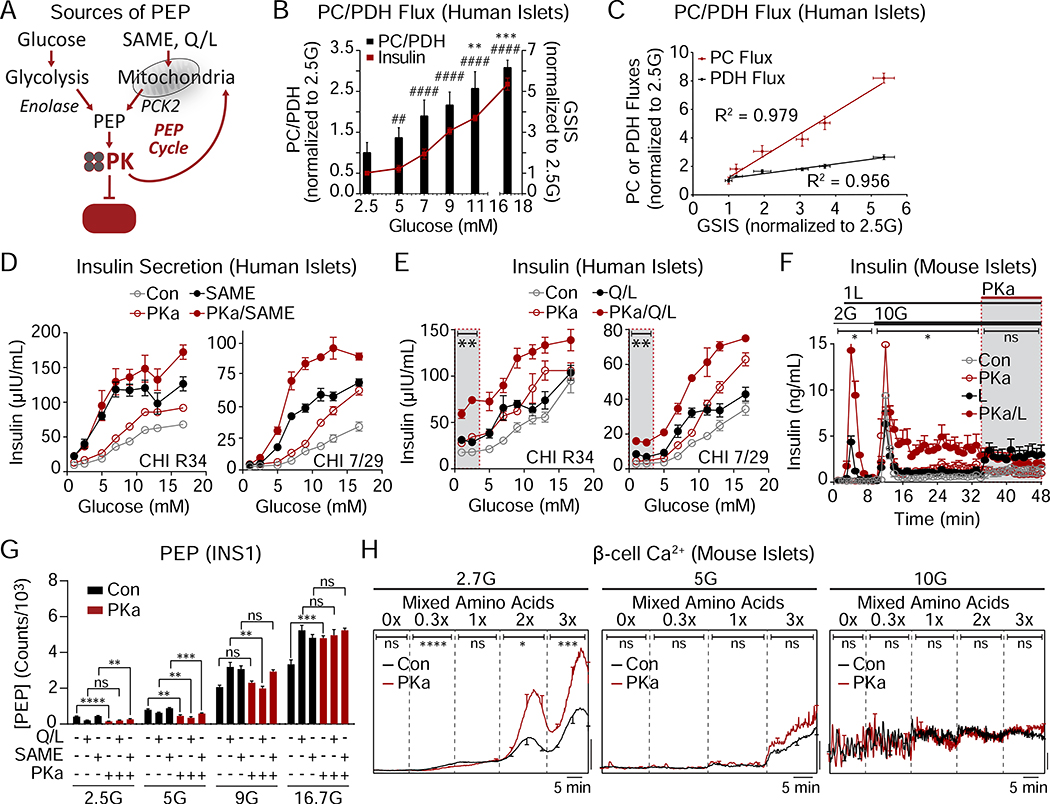
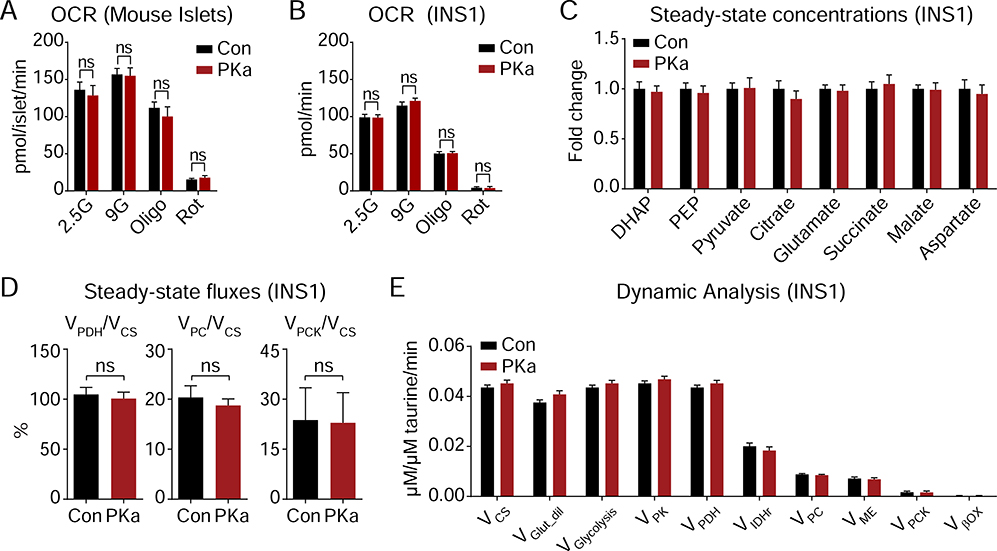
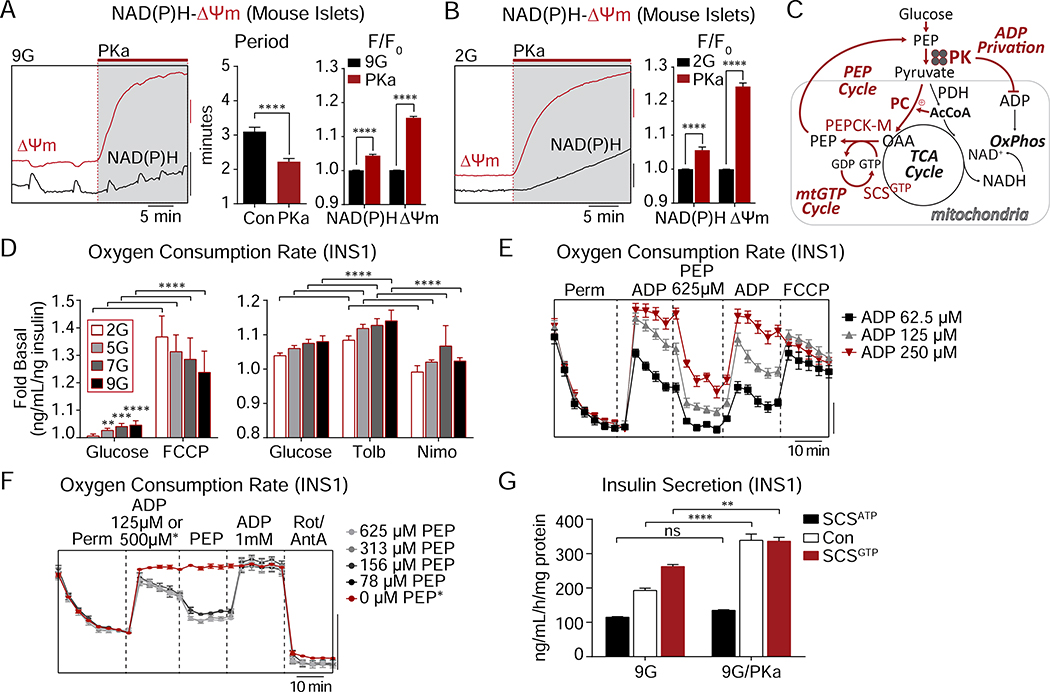
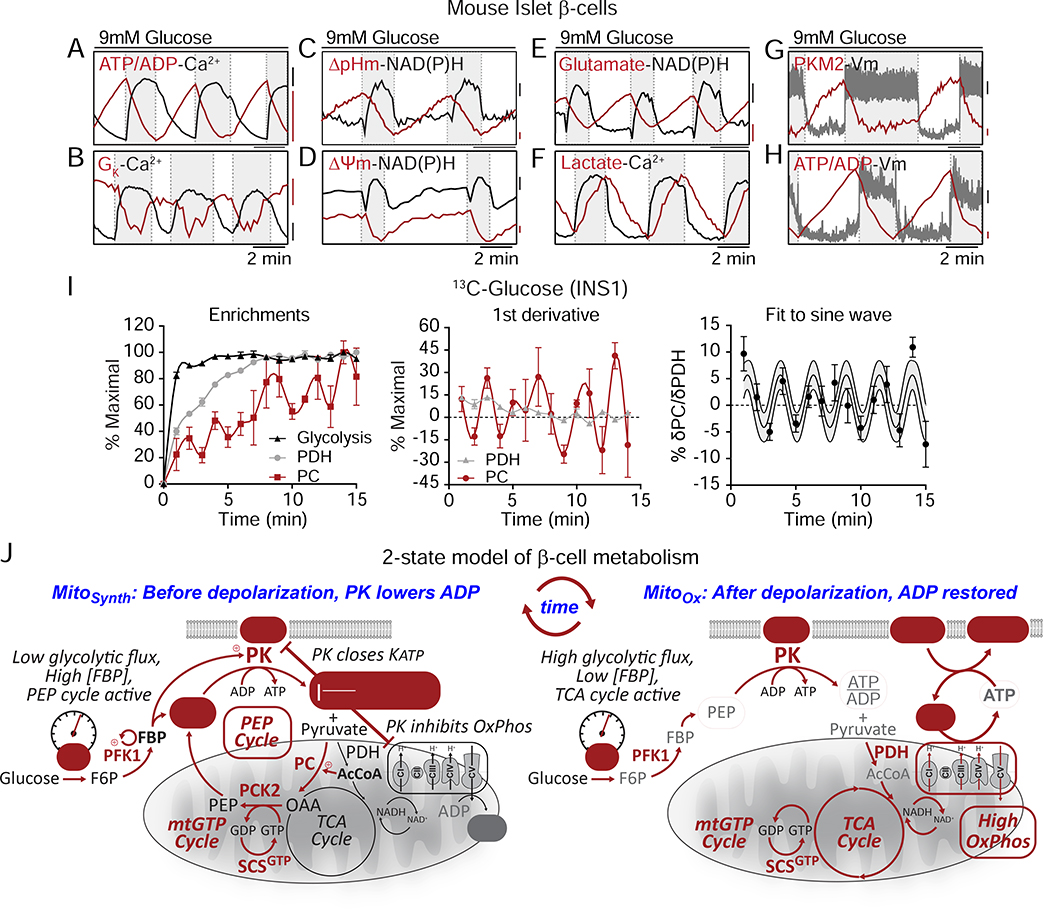
Pancreatic β cells couple nutrient metabolism with appropriate insulin secretion. Here, we show that pyruvate kinase (PK), which converts ADP and phosphoenolpyruvate (PEP) into ATP and pyruvate, underlies β cell sensing of both glycolytic and mitochondrial fuels. Plasma membrane-localized PK is sufficient to close KATP channels and initiate calcium influx. Small-molecule PK activators increase the frequency of ATP/ADP and calcium oscillations and potently amplify insulin secretion. PK restricts respiration by cyclically depriving mitochondria of ADP, which accelerates PEP cycling until membrane depolarization restores ADP and oxidative phosphorylation. Our findings support a compartmentalized model of β cell metabolism in which PK locally generates the ATP/ADP required for insulin secretion. Oscillatory PK activity allows mitochondria to perform synthetic and oxidative functions without any net impact on glucose oxidation. These findings suggest a potential therapeutic route for diabetes based on PK activation that would not be predicted by the current consensus single-state model of β cell function.
Keywords: K(ATP) channel; anaplerosis; biosensor imaging; insulin secretion; metabolic flux; metabolic oscillations; oxidative phosphorylation; phosphoenolpyruvate cycle; pyruvate kinase; β cell metabolism.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
Comment in
-
Targeting Pyruvate Kinase PEPs Up Insulin Secretion and Improves Glucose Homeostasis.Cell Metab. 2020 Nov 3;32(5):693-694. doi: 10.1016/j.cmet.2020.10.008. Cell Metab. 2020. PMID: 33147479
-
Pyruvate kinase regulates insulin secretion.Nat Rev Endocrinol. 2021 Jan;17(1):3. doi: 10.1038/s41574-020-00447-0. Nat Rev Endocrinol. 2021. PMID: 33208920 No abstract available.
References
-
- Abulizi A, Stark R, Cardone RL, Lewandowski SL, Zhao X, Alves TC, Thomas C, Kung C, Wang B, Siebel S, et al. (2020). Pharmacologic activation of the mitochondrial phosphoenolpyruvate cycle enhances islet function in vivo. BioRxiv 2020.02.13.947630.
-
- Affourtit C, Alberts B, Barlow J, Carré JE, and Wynne AG (2018). Control of pancreatic β-cell bioenergetics. Biochem. Soc. Trans. 46, 555–564. - PubMed
-
- Ainscow EK, and Rutter GA (2002). Glucose-stimulated oscillations in free cytosolic ATP concentration imaged in single islet beta-cells: evidence for a Ca2+-dependent mechanism. Diabetes 51 Suppl 1, S162–170. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 DK110324/DK/NIDDK NIH HHS/United States
- R01 DK113103/DK/NIDDK NIH HHS/United States
- R21 AG050135/AG/NIA NIH HHS/United States
- UL1 TR001863/TR/NCATS NIH HHS/United States
- F32 DK116542/DK/NIDDK NIH HHS/United States
- R01 DK110181/DK/NIDDK NIH HHS/United States
- I01 BX004715/BX/BLRD VA/United States
- T32 DK007665/DK/NIDDK NIH HHS/United States
- K08 DK080142/DK/NIDDK NIH HHS/United States
- F31 DK126403/DK/NIDDK NIH HHS/United States
- UL1 RR024139/RR/NCRR NIH HHS/United States
- K01 DK101683/DK/NIDDK NIH HHS/United States
- R01 AG062328/AG/NIA NIH HHS/United States
- R01 DK127637/DK/NIDDK NIH HHS/United States
- T32 AG000213/AG/NIA NIH HHS/United States
- P30 DK045735/DK/NIDDK NIH HHS/United States
- R01 DK092606/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
