

Multi-Tissue Acceleration of the Mitochondrial Phosphoenolpyruvate Cycle Improves Whole-Body Metabolic Health
- PMID: 33147485
- PMCID: PMC7679013
- DOI: 10.1016/j.cmet.2020.10.006
Multi-Tissue Acceleration of the Mitochondrial Phosphoenolpyruvate Cycle Improves Whole-Body Metabolic Health
Abstract
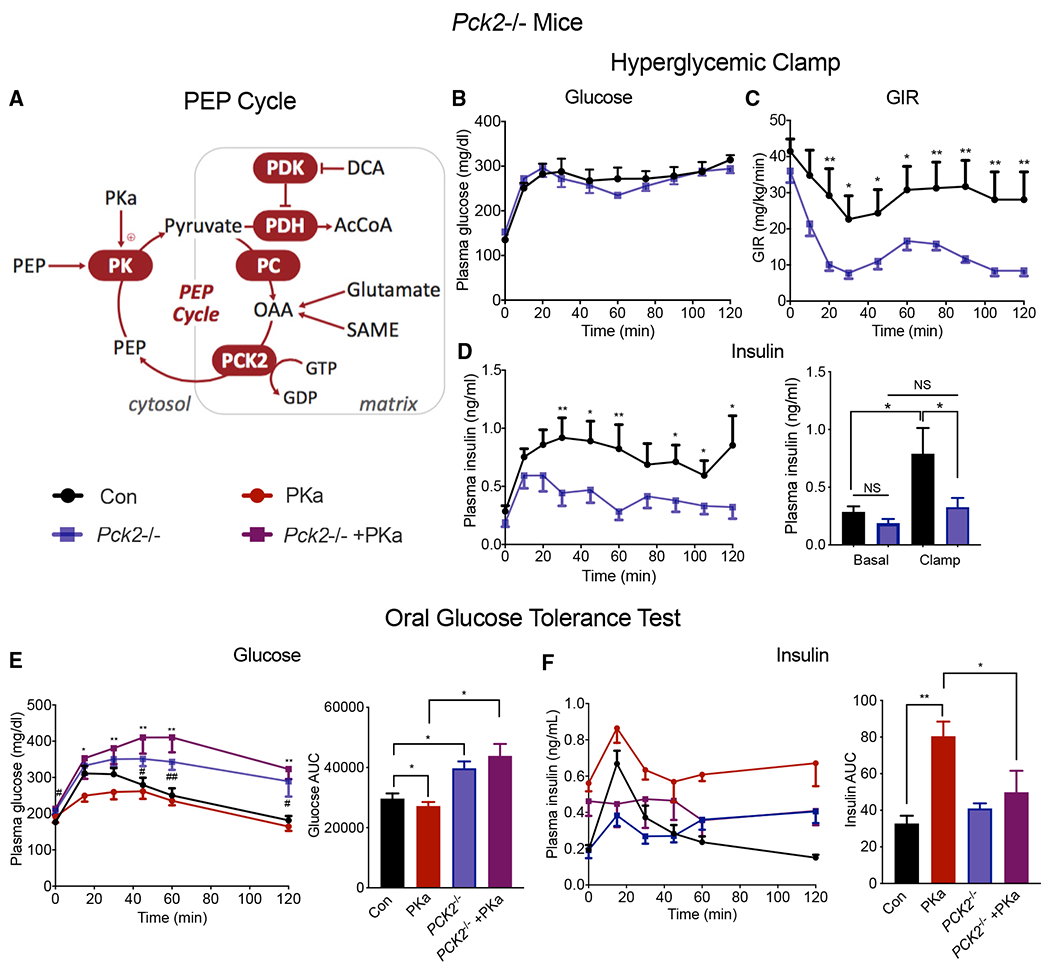
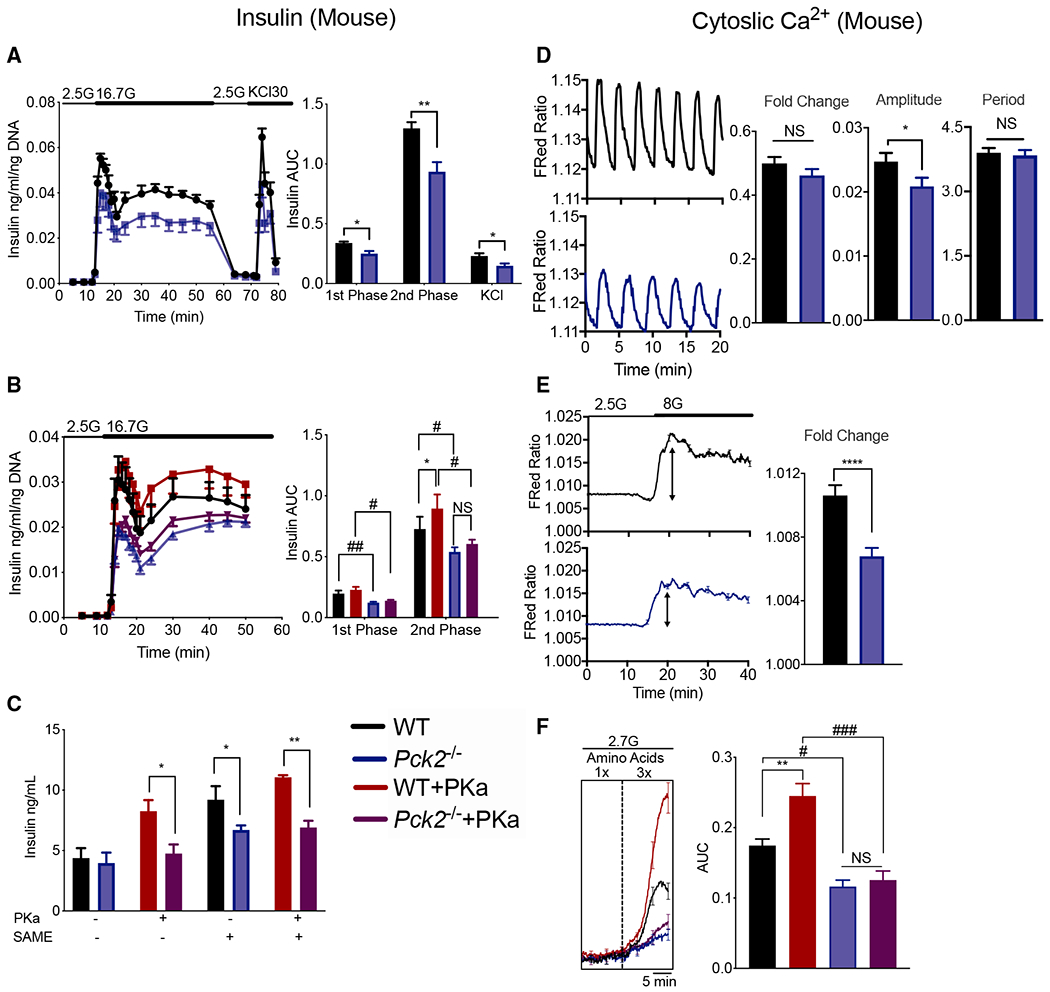
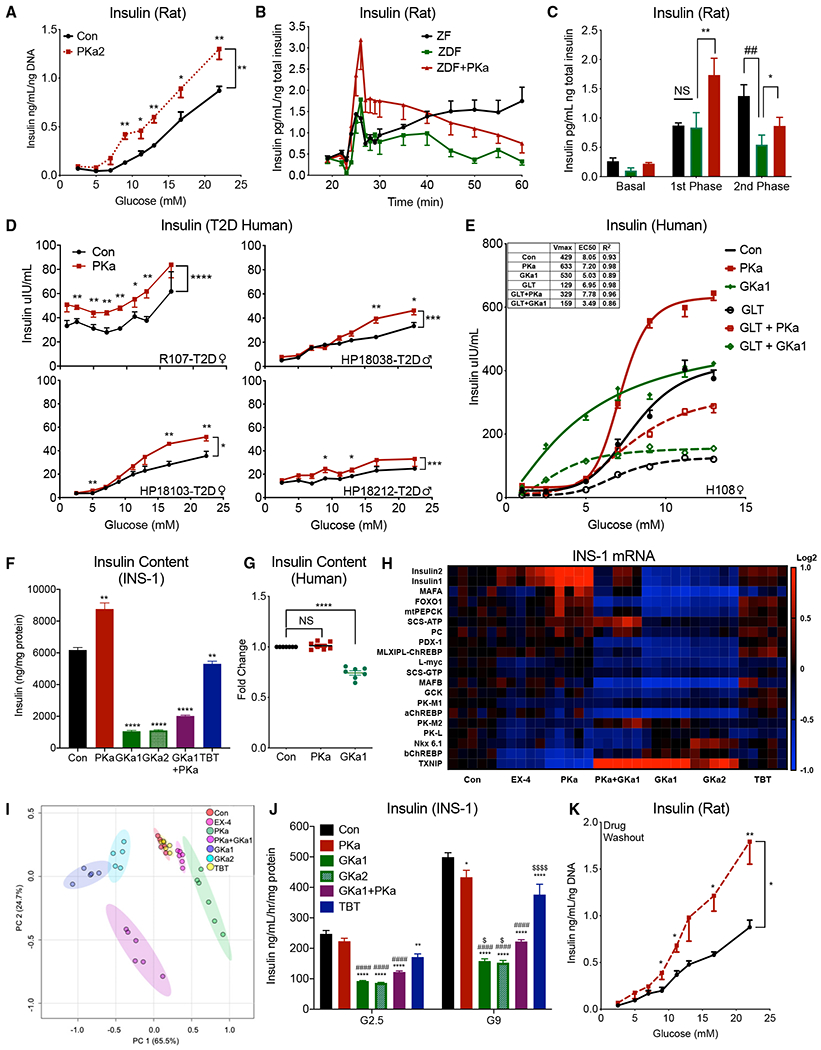
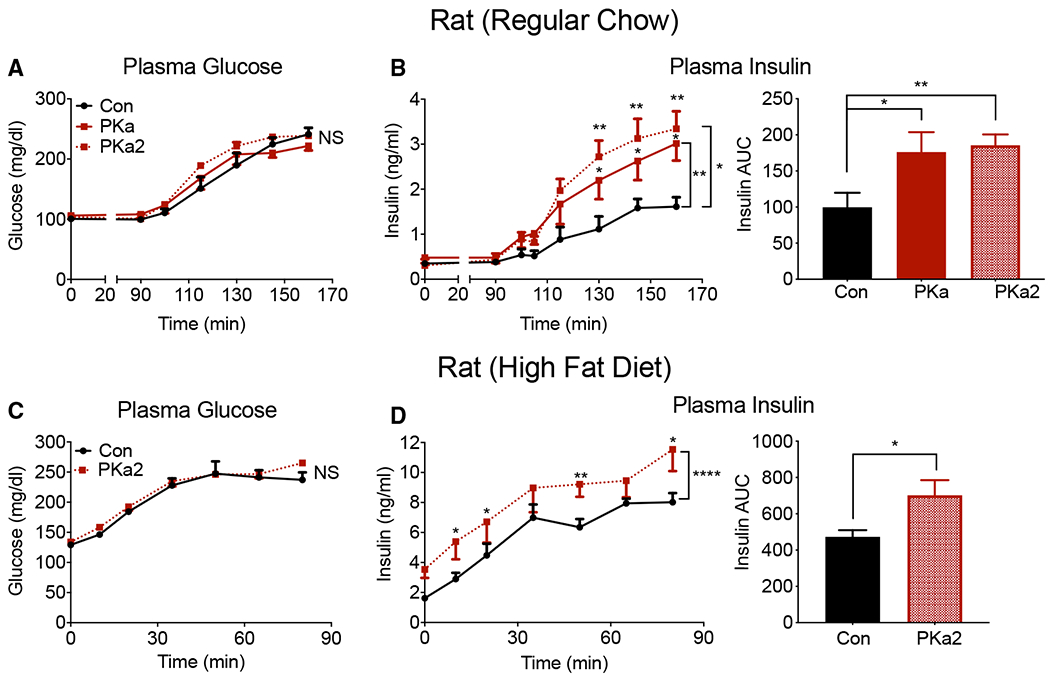
The mitochondrial GTP (mtGTP)-dependent phosphoenolpyruvate (PEP) cycle couples mitochondrial PEPCK (PCK2) to pyruvate kinase (PK) in the liver and pancreatic islets to regulate glucose homeostasis. Here, small molecule PK activators accelerated the PEP cycle to improve islet function, as well as metabolic homeostasis, in preclinical rodent models of diabetes. In contrast, treatment with a PK activator did not improve insulin secretion in pck2-/- mice. Unlike other clinical secretagogues, PK activation enhanced insulin secretion but also had higher insulin content and markers of differentiation. In addition to improving insulin secretion, acute PK activation short-circuited gluconeogenesis to reduce endogenous glucose production while accelerating red blood cell glucose turnover. Four-week delivery of a PK activator in vivo remodeled PK phosphorylation, reduced liver fat, and improved hepatic and peripheral insulin sensitivity in HFD-fed rats. These data provide a preclinical rationale for PK activation to accelerate the PEP cycle to improve metabolic homeostasis and insulin sensitivity.
Keywords: anaplerosis; cataplerosis; fatty liver; human islets; insulin resistance; insulin secretion; mitochondrial GTP; mitochondrial PEPCK; phosphoenolpyruvate cycle; pyruvate kinase.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests C.K. is an employee of and stockholder in Agios Pharmaceuticals. R.G.K. has been a paid consultant for Agios Pharmaceuticals. The remaining authors declare no competing interests.
Figures

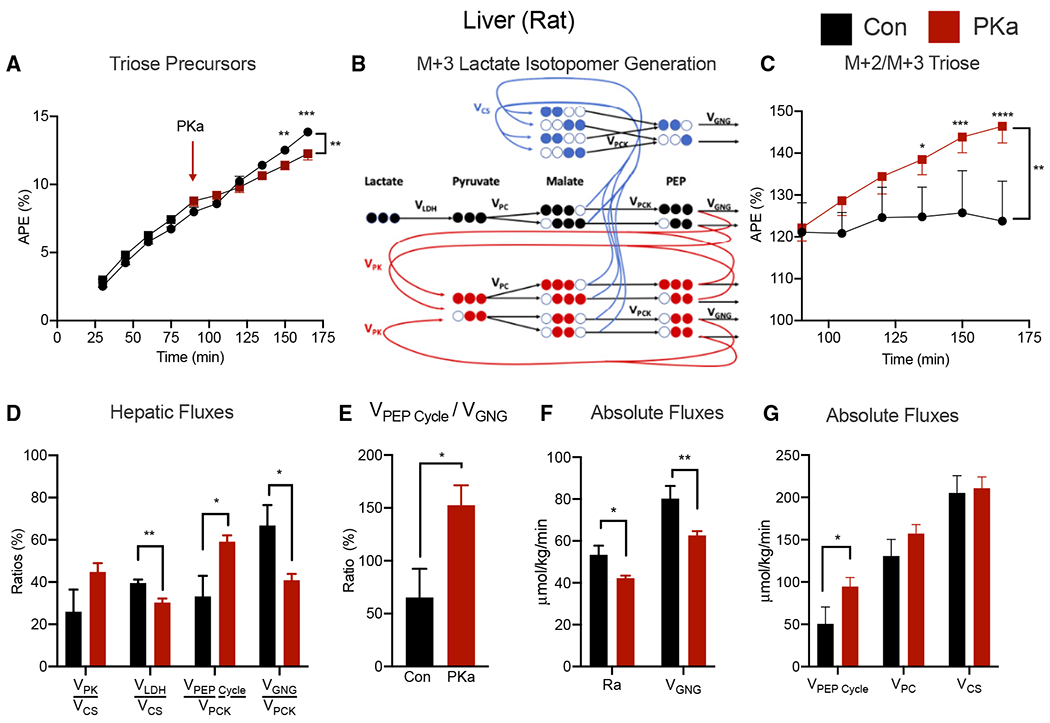
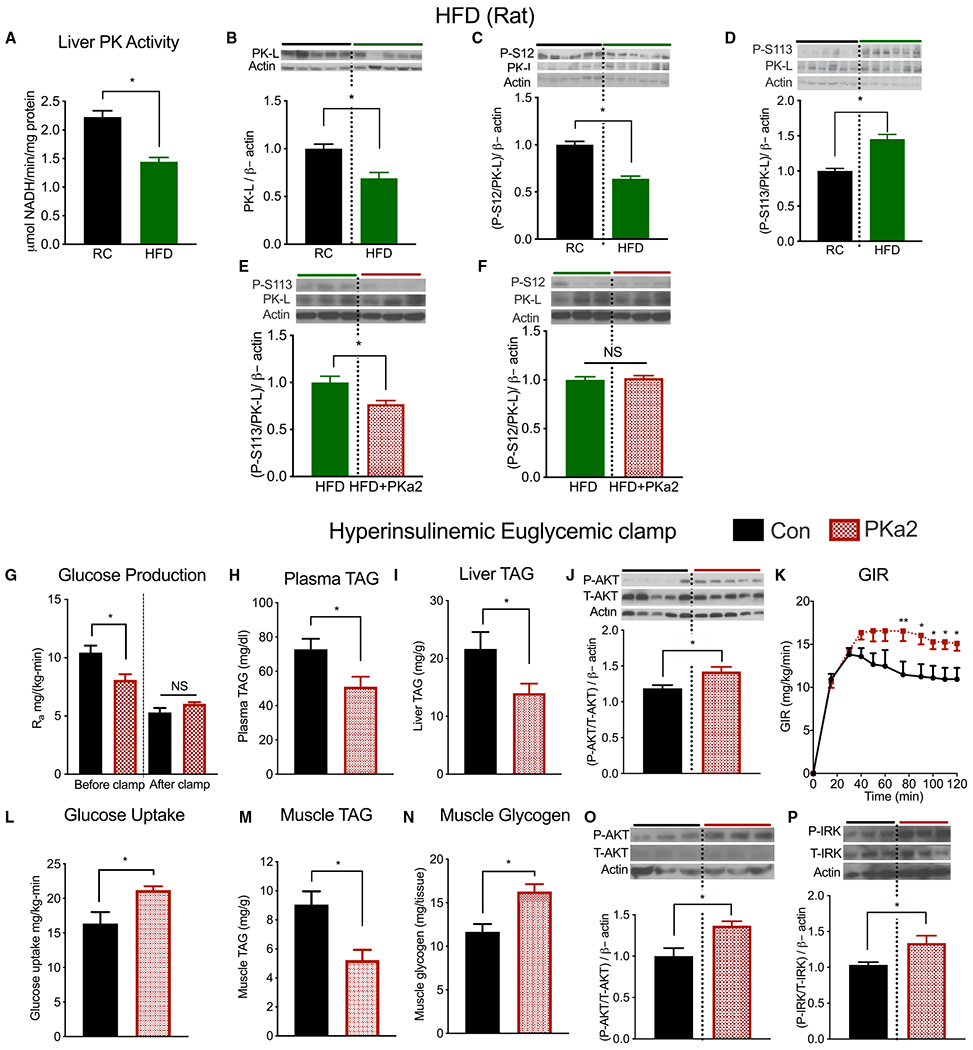
Comment in
-
Targeting Pyruvate Kinase PEPs Up Insulin Secretion and Improves Glucose Homeostasis.Cell Metab. 2020 Nov 3;32(5):693-694. doi: 10.1016/j.cmet.2020.10.008. Cell Metab. 2020. PMID: 33147479
-
Pyruvate kinase regulates insulin secretion.Nat Rev Endocrinol. 2021 Jan;17(1):3. doi: 10.1038/s41574-020-00447-0. Nat Rev Endocrinol. 2021. PMID: 33208920 No abstract available.
References
-
- Akhmedov D, De Marchi U, Wollheim CB, and Wiederkehr A (2012). Pyruvate dehydrogenase E1α phosphorylation is induced by glucose but does not control metabolism-secretion coupling in INS-1E clonal β-cells. Biochim. Biophys. Acta 1823, 1815–1824. - PubMed
-
- Cline GW, Lepine RL, Papas KK, Kibbey RG, and Shulman GI (2004). 13C NMR isotopomer analysis of anaplerotic pathways in INS-1 cells. J. Biol. Chem 279, 44370–44375. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 DK113103/DK/NIDDK NIH HHS/United States
- R21 AG050135/AG/NIA NIH HHS/United States
- UL1 TR001863/TR/NCATS NIH HHS/United States
- R01 DK110181/DK/NIDDK NIH HHS/United States
- R01 DK127637/DK/NIDDK NIH HHS/United States
- T32 DK007665/DK/NIDDK NIH HHS/United States
- P30 DK034989/DK/NIDDK NIH HHS/United States
- K08 DK080142/DK/NIDDK NIH HHS/United States
- F31 DK126403/DK/NIDDK NIH HHS/United States
- UL1 RR024139/RR/NCRR NIH HHS/United States
- R01 DK108283/DK/NIDDK NIH HHS/United States
- K01 DK101683/DK/NIDDK NIH HHS/United States
- R01 AG062328/AG/NIA NIH HHS/United States
- P30 DK045735/DK/NIDDK NIH HHS/United States
- R01 DK092606/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
