

Diversity of Long-Lived Intermediates along the Binding Pathway of Imatinib to Abl Kinase Revealed by MD Simulations
- PMID: 33147951
- PMCID: PMC8288498
- DOI: 10.1021/acs.jctc.0c00739
Diversity of Long-Lived Intermediates along the Binding Pathway of Imatinib to Abl Kinase Revealed by MD Simulations
Abstract
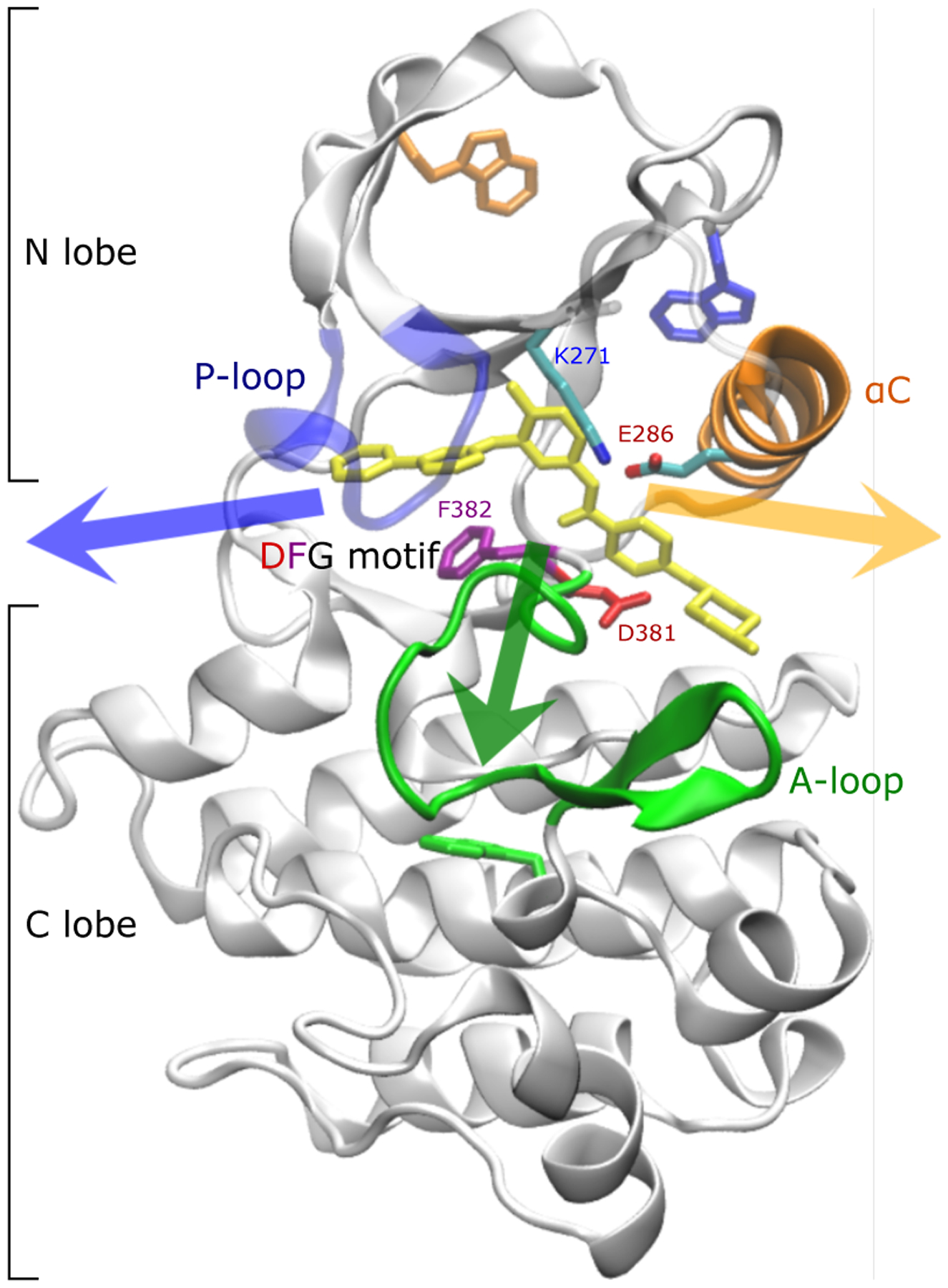
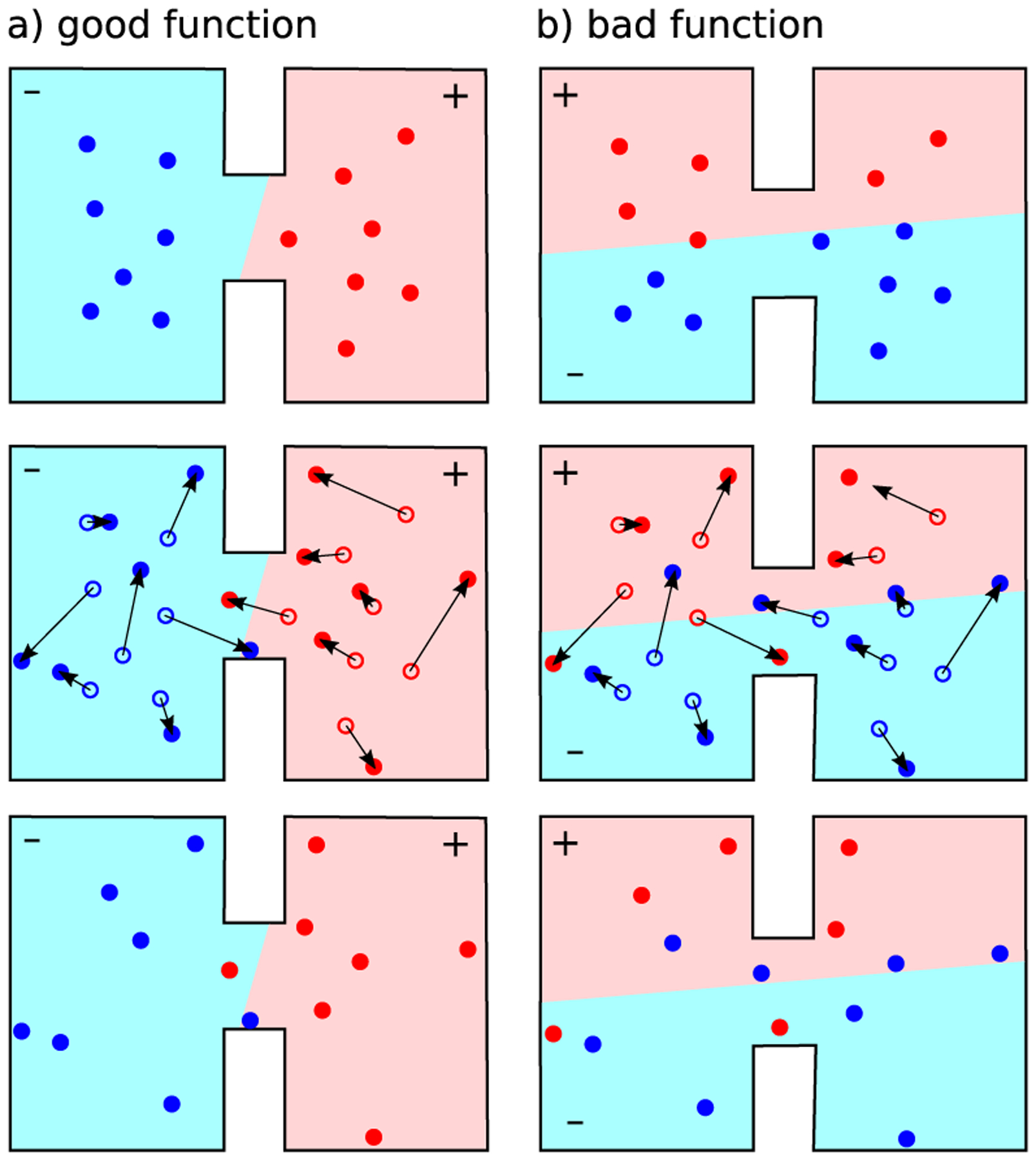
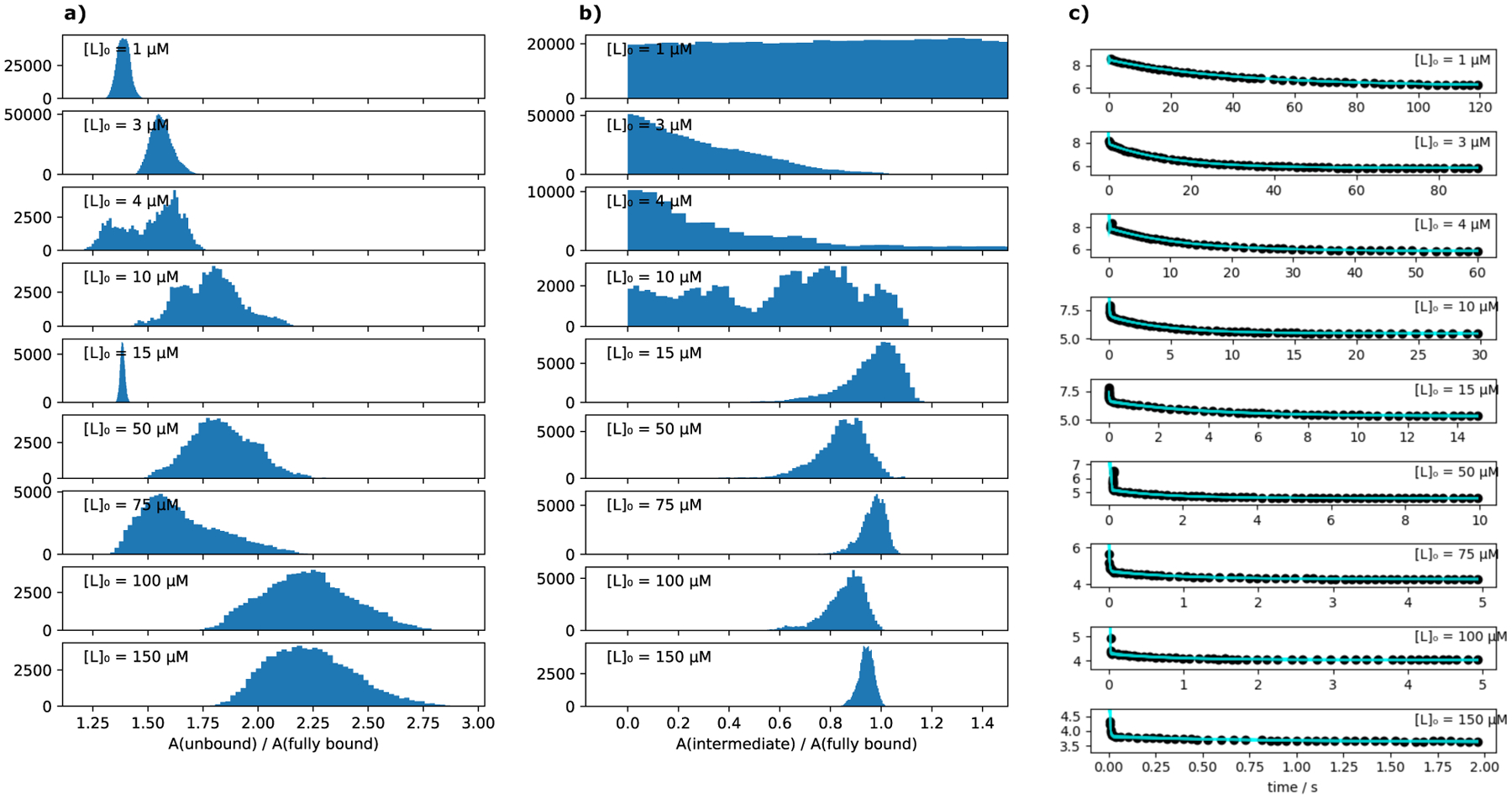
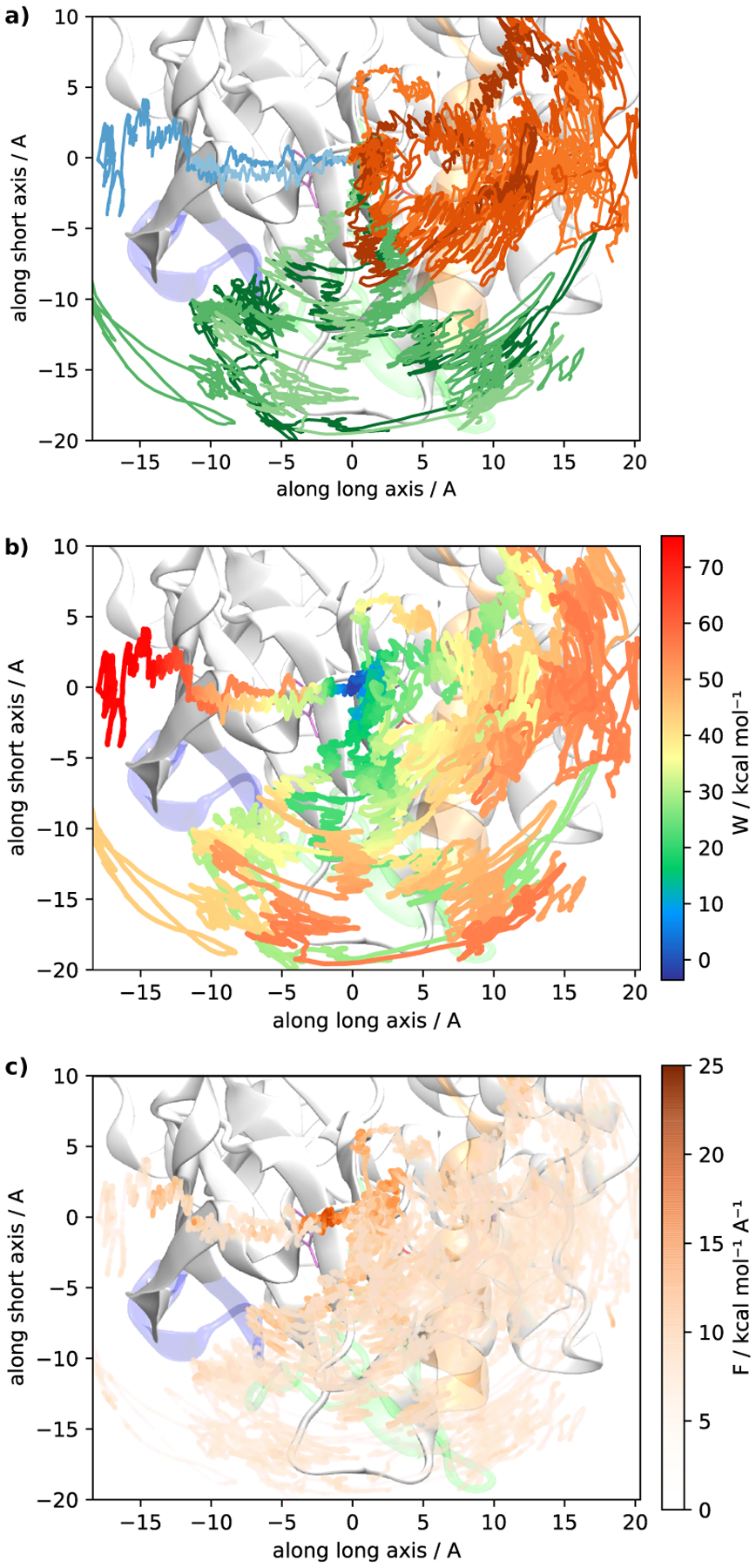
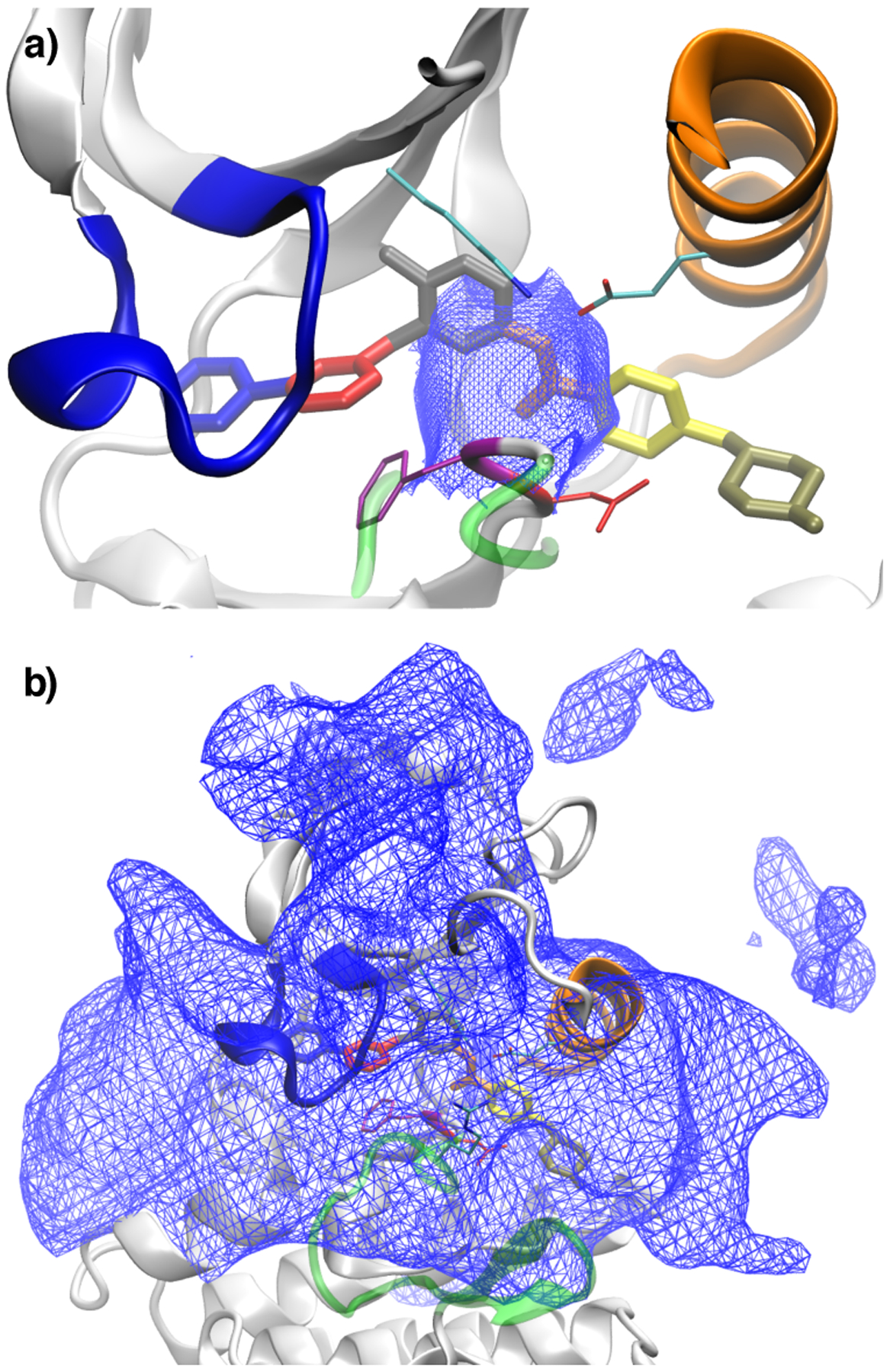
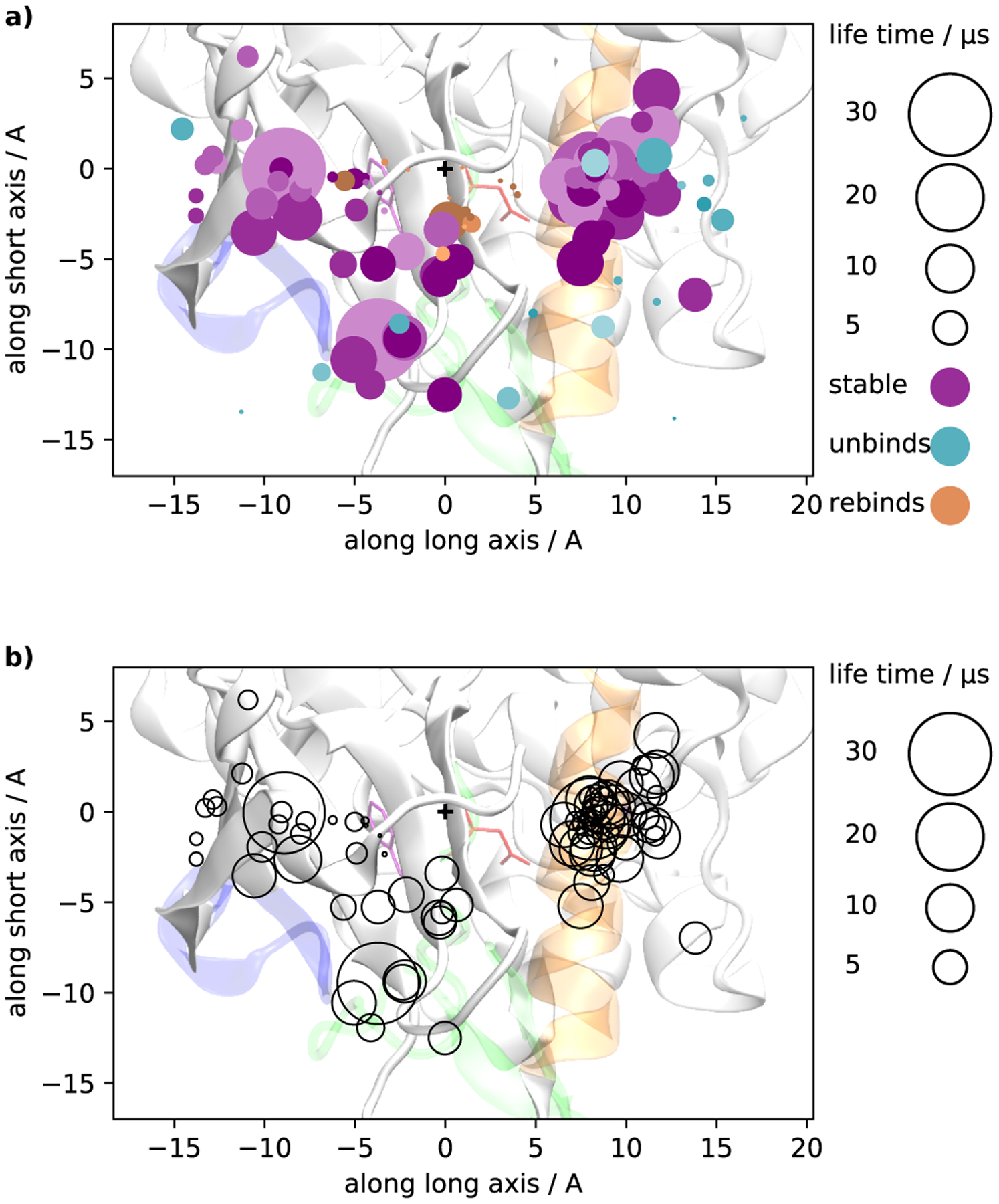
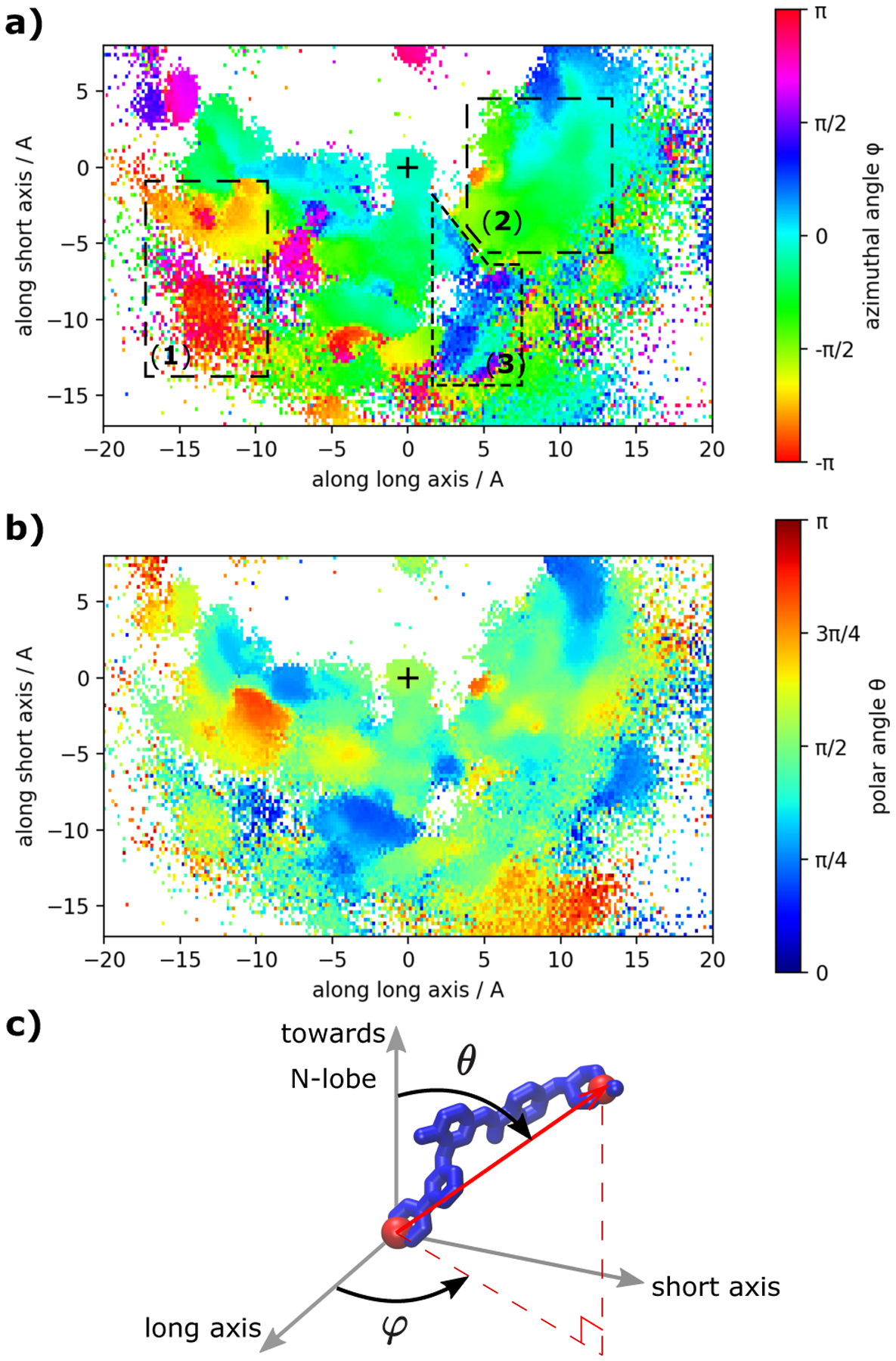
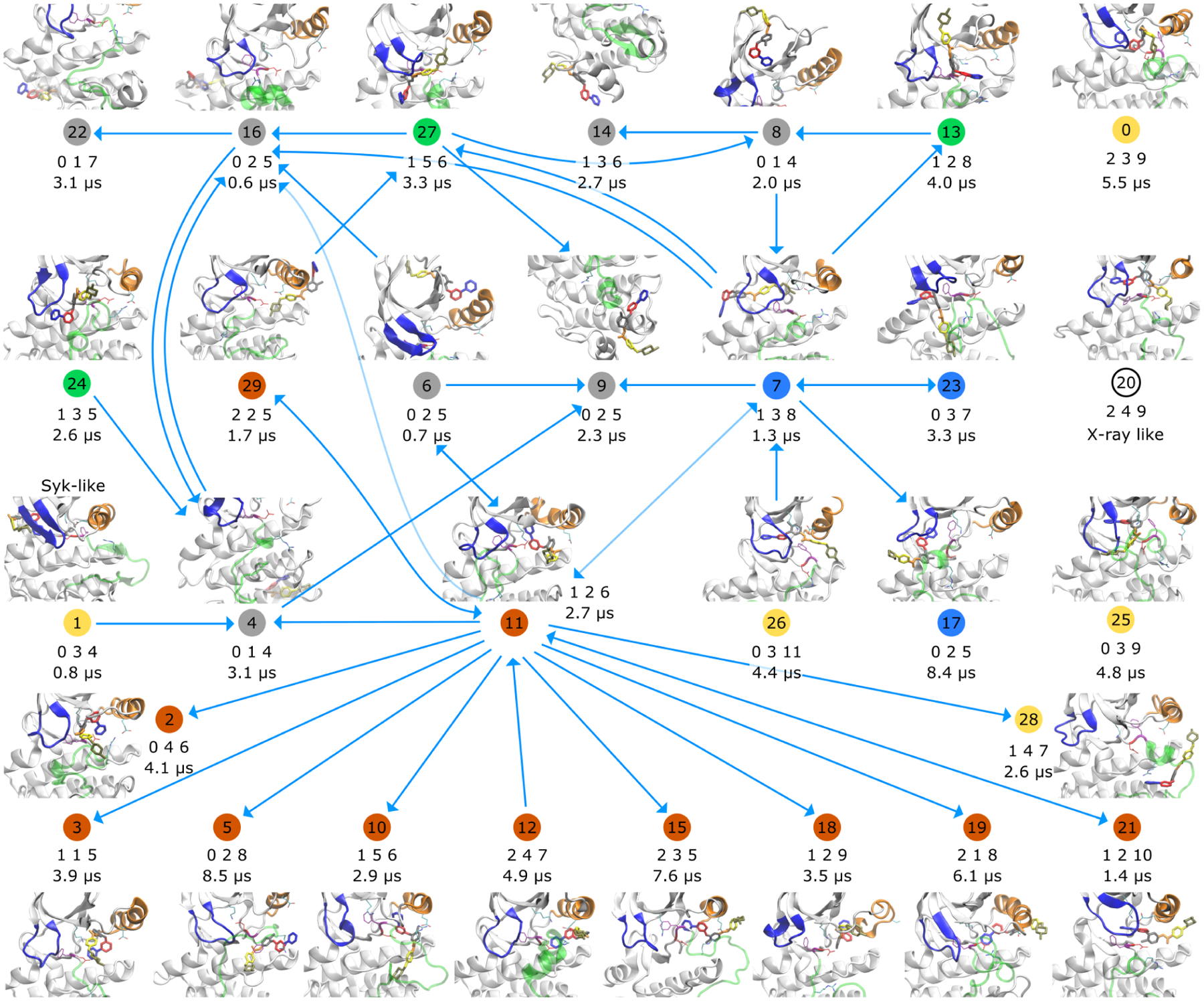
Imatinib, a drug used for the treatment of chronic myeloid leukemia and other cancers, works by blocking the catalytic site of pathological constitutively active Abl kinase. While the binding pose is known from X-ray crystallography, the different steps leading to the formation of the complex are not well understood. The results from extensive molecular dynamics simulations show that imatinib can primarily exit the known crystallographic binding pose through the cleft of the binding site or by sliding under the αC helix. Once displaced from the crystallographic binding pose, imatinib becomes trapped in intermediate states. These intermediates are characterized by a high diversity of ligand orientations and conformations, and relaxation timescales within this region may exceed 3-4 ms. Analysis indicates that the metastable intermediate states should be spectroscopically indistinguishable from the crystallographic binding pose, in agreement with tryptophan stopped-flow fluorescence experiments.
Figures
Similar articles
-
Drug repurposing for chronic myeloid leukemia: in silico and in vitro investigation of DrugBank database for allosteric Bcr-Abl inhibitors.J Biomol Struct Dyn. 2017 Jun;35(8):1833-1848. doi: 10.1080/07391102.2016.1196462. Epub 2016 Jun 29. J Biomol Struct Dyn. 2017. PMID: 27353341
-
Dual-specific Src and Abl kinase inhibitors, PP1 and CGP76030, inhibit growth and survival of cells expressing imatinib mesylate-resistant Bcr-Abl kinases.Blood. 2003 Jan 15;101(2):664-72. doi: 10.1182/blood-2002-01-0288. Epub 2002 Sep 5. Blood. 2003. PMID: 12393636
-
Molecular dynamics simulations provide insights into the origin of gleevec's selectivity toward human tyrosine kinases.J Biomol Struct Dyn. 2019 Jul;37(10):2733-2744. doi: 10.1080/07391102.2018.1496139. Epub 2018 Nov 1. J Biomol Struct Dyn. 2019. PMID: 30052122
-
[MET/ERK and MET/JNK Pathway Activation Is Involved in BCR-ABL Inhibitor-resistance in Chronic Myeloid Leukemia].Yakugaku Zasshi. 2018;138(12):1461-1466. doi: 10.1248/yakushi.18-00142. Yakugaku Zasshi. 2018. PMID: 30504658 Review. Japanese.
-
Last findings on dual inhibitors of abl and SRC tyrosine-kinases.Mini Rev Med Chem. 2007 Feb;7(2):191-201. doi: 10.2174/138955707779802598. Mini Rev Med Chem. 2007. PMID: 17305593 Review.
Cited by
-
A Guide to In Silico Drug Design.Pharmaceutics. 2022 Dec 23;15(1):49. doi: 10.3390/pharmaceutics15010049. Pharmaceutics. 2022. PMID: 36678678 Free PMC article. Review.
-
Integration of a Randomized Sequence Scanning Approach in AlphaFold2 and Local Frustration Profiling of Conformational States Enable Interpretable Atomistic Characterization of Conformational Ensembles and Detection of Hidden Allosteric States in the ABL1 Protein Kinase.J Chem Theory Comput. 2024 Jun 25;20(12):5317-5336. doi: 10.1021/acs.jctc.4c00222. Epub 2024 Jun 12. J Chem Theory Comput. 2024. PMID: 38865109 Free PMC article.
-
Markov State Models: To Optimize or Not to Optimize.J Chem Theory Comput. 2024 Jan 23;20(2):977-988. doi: 10.1021/acs.jctc.3c01134. Epub 2024 Jan 1. J Chem Theory Comput. 2024. PMID: 38163961 Free PMC article.
-
Treatment of flexibility of protein backbone in simulations of protein-ligand interactions using steered molecular dynamics.Sci Rep. 2024 May 7;14(1):10475. doi: 10.1038/s41598-024-59899-3. Sci Rep. 2024. PMID: 38714683 Free PMC article.
-
Predicting Mutation-Induced Allosteric Changes in Structures and Conformational Ensembles of the ABL Kinase Using AlphaFold2 Adaptations with Alanine Sequence Scanning.Int J Mol Sci. 2024 Sep 19;25(18):10082. doi: 10.3390/ijms251810082. Int J Mol Sci. 2024. PMID: 39337567 Free PMC article.
References
-
- Druker BJ; Tamura S; Buchdunger E; Ohno S; Segal GM; Fanning S; Zimmermann J; Lydon NB Effects of a selective inhibitor of the Abl tyrosine kinase on the growth of Bcr-Abl positive cells. Nat. Med 1996, 2, 561–566. - PubMed
-
- Mauro MJ; Druker BJ STI571: targeting BCR-Abl as therapy for CML. Oncologist 2001, 6, 233–238. - PubMed
-
- Melo J The diversity of BCR-ABL fusion proteins and their relationship to leukemia phenotype. Blood 1996, 88, 2375–2384. - PubMed
-
- Nagar B; Bornmann WG; Pellicena P; Schindler T; Veach DR; Miller WT; Clarkson B; Kuriyan J Crystal Structures of the Kinase Domain of c-Abl in Complex with the Small Molecule Inhibitors PD173955 and Imatinib (STI-571). Cancer Res. 2002, 62, 4236–4243. - PubMed
-
- Roskoski R A historical overview of protein kinases and their targeted small molecule inhibitors. Pharmacol. Res 2015, 100, 1–23. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous