

Small-molecule sequestration of amyloid-β as a drug discovery strategy for Alzheimer's disease
- PMID: 33148639
- PMCID: PMC7673680
- DOI: 10.1126/sciadv.abb5924
Small-molecule sequestration of amyloid-β as a drug discovery strategy for Alzheimer's disease
Abstract
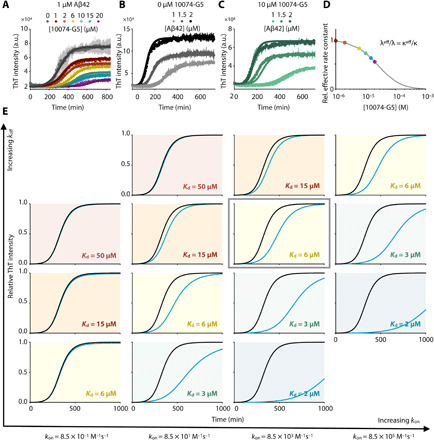
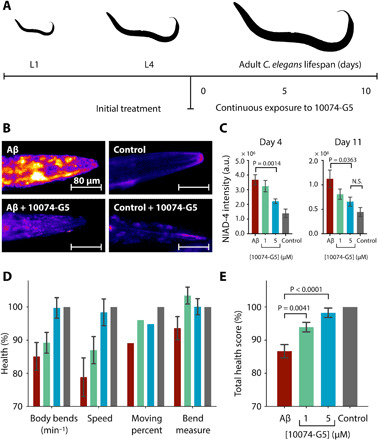
Disordered proteins are challenging therapeutic targets, and no drug is currently in clinical use that modifies the properties of their monomeric states. Here, we identify a small molecule (10074-G5) capable of binding and sequestering the intrinsically disordered amyloid-β (Aβ) peptide in its monomeric, soluble state. Our analysis reveals that this compound interacts with Aβ and inhibits both the primary and secondary nucleation pathways in its aggregation process. We characterize this interaction using biophysical experiments and integrative structural ensemble determination methods. We observe that this molecule increases the conformational entropy of monomeric Aβ while decreasing its hydrophobic surface area. We also show that it rescues a Caenorhabditis elegans model of Aβ-associated toxicity, consistent with the mechanism of action identified from the in silico and in vitro studies. These results illustrate the strategy of stabilizing the monomeric states of disordered proteins with small molecules to alter their behavior for therapeutic purposes.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution License 4.0 (CC BY).
Figures
Similar articles
-
Small-Molecule Targeted Aβ42 Aggregate Degradation: Negatively Charged Small Molecules Are More Promising than the Neutral Ones.ACS Chem Neurosci. 2021 Apr 7;12(7):1197-1209. doi: 10.1021/acschemneuro.1c00047. Epub 2021 Mar 9. ACS Chem Neurosci. 2021. PMID: 33687193
-
A Small Molecule Stabilizes the Disordered Native State of the Alzheimer's Aβ Peptide.ACS Chem Neurosci. 2022 Jun 15;13(12):1738-1745. doi: 10.1021/acschemneuro.2c00116. Epub 2022 Jun 1. ACS Chem Neurosci. 2022. PMID: 35649268 Free PMC article.
-
The inhibitory mechanism of a fullerene derivative against amyloid-β peptide aggregation: an atomistic simulation study.Phys Chem Chem Phys. 2016 May 14;18(18):12582-91. doi: 10.1039/c6cp01014h. Epub 2016 Apr 19. Phys Chem Chem Phys. 2016. PMID: 27091578
-
An Overview of Several Inhibitors for Alzheimer's Disease: Characterization and Failure.Int J Mol Sci. 2021 Oct 6;22(19):10798. doi: 10.3390/ijms221910798. Int J Mol Sci. 2021. PMID: 34639140 Free PMC article. Review.
-
An overview of glutaminyl cyclase inhibitors for Alzheimer's disease.Future Med Chem. 2019 Dec;11(24):3179-3194. doi: 10.4155/fmc-2019-0163. Future Med Chem. 2019. PMID: 31838899 Review.
Cited by
-
Advances in targeting 'undruggable' transcription factors with small molecules.Nat Rev Drug Discov. 2021 Sep;20(9):669-688. doi: 10.1038/s41573-021-00199-0. Epub 2021 May 18. Nat Rev Drug Discov. 2021. PMID: 34006959 Review.
-
Molecular Mechanisms of Inhibition of Protein Amyloid Fibril Formation: Evidence and Perspectives Based on Kinetic Models.Int J Mol Sci. 2022 Nov 3;23(21):13428. doi: 10.3390/ijms232113428. Int J Mol Sci. 2022. PMID: 36362217 Free PMC article. Review.
-
Molecular simulations integrated with experiments for probing the interaction dynamics and binding mechanisms of intrinsically disordered proteins.Curr Opin Struct Biol. 2024 Feb;84:102756. doi: 10.1016/j.sbi.2023.102756. Epub 2023 Dec 19. Curr Opin Struct Biol. 2024. PMID: 38118365 Free PMC article. Review.
-
Kinetic Steering of Amyloid Formation and Polymorphism by Canagliflozin, a Type-2 Diabetes Drug.J Am Chem Soc. 2025 Apr 9;147(14):11859-11878. doi: 10.1021/jacs.4c16743. Epub 2025 Feb 21. J Am Chem Soc. 2025. PMID: 39985130 Free PMC article.
-
Evidence of α-Synuclein/Glucocerebrosidase Dual Targeting by Iminosugar Derivatives.ACS Chem Neurosci. 2025 Apr 2;16(7):1251-1257. doi: 10.1021/acschemneuro.4c00618. Epub 2025 Mar 13. ACS Chem Neurosci. 2025. PMID: 40079830 Free PMC article.
References
-
- Knowles T. P., Vendruscolo M., Dobson C. M., The amyloid state and its association with protein misfolding diseases. Nat. Rev. Mol. Cell Biol. 15, 384–396 (2014). - PubMed
-
- Hardy J., Selkoe D. J., The amyloid hypothesis of Alzheimer’s disease: Progress and problems on the road to therapeutics. Science 297, 353–356 (2002). - PubMed
-
- Haass C., Selkoe D. J., Soluble protein oligomers in neurodegeneration: Lessons from the Alzheimer’s amyloid β-peptide. Nat. Rev. Mol. Cell Biol. 8, 101–112 (2007). - PubMed
