

Epigallocatechin Gallate Effectively Affects Senescence and Anti-SASP via SIRT3 in 3T3-L1 Preadipocytes in Comparison with Other Bioactive Substances
- PMID: 33149809
- PMCID: PMC7603628
- DOI: 10.1155/2020/4793125
Epigallocatechin Gallate Effectively Affects Senescence and Anti-SASP via SIRT3 in 3T3-L1 Preadipocytes in Comparison with Other Bioactive Substances
Abstract
Aim: We investigated different bioactive compounds including epigallocatechin gallate (EGCG), anthocyanidin, resveratrol, phloretin, spermidine, butyrate, and β-hydroxybutyrate with regard to their effect on SIRT3 via NRF2 and modulation of the proinflammatory senescence-associated secretory phenotype (SASP) in senescence induced 3T3-L1 preadipocytes.
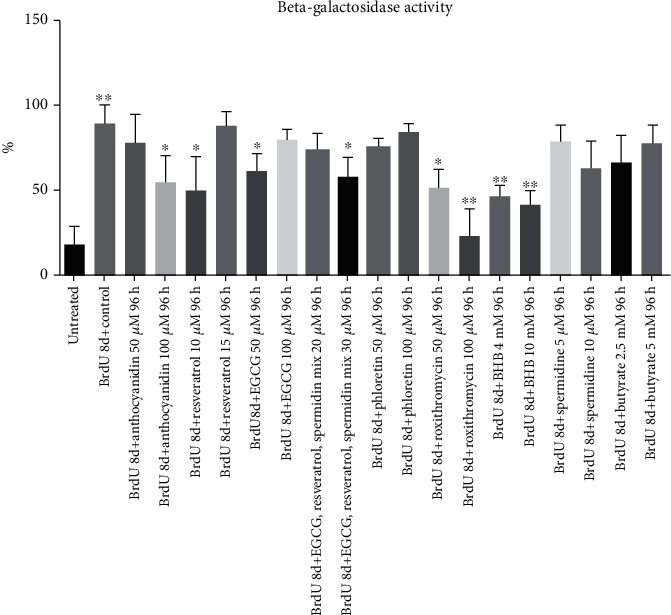
Methods: For induction of senescence, 3T3-L1 preadipocytes were incubated with bromodeoxyuridine (BrdU) for 8 days. Cell cycle inhibition was observed, and β-galactosidase activity was measured. After BrdU treatment, cells were treated with different bioactive compounds in various concentrations for 96 h. ELISA was used for determining proinflammatory cytokine IL6 in SASP cells.
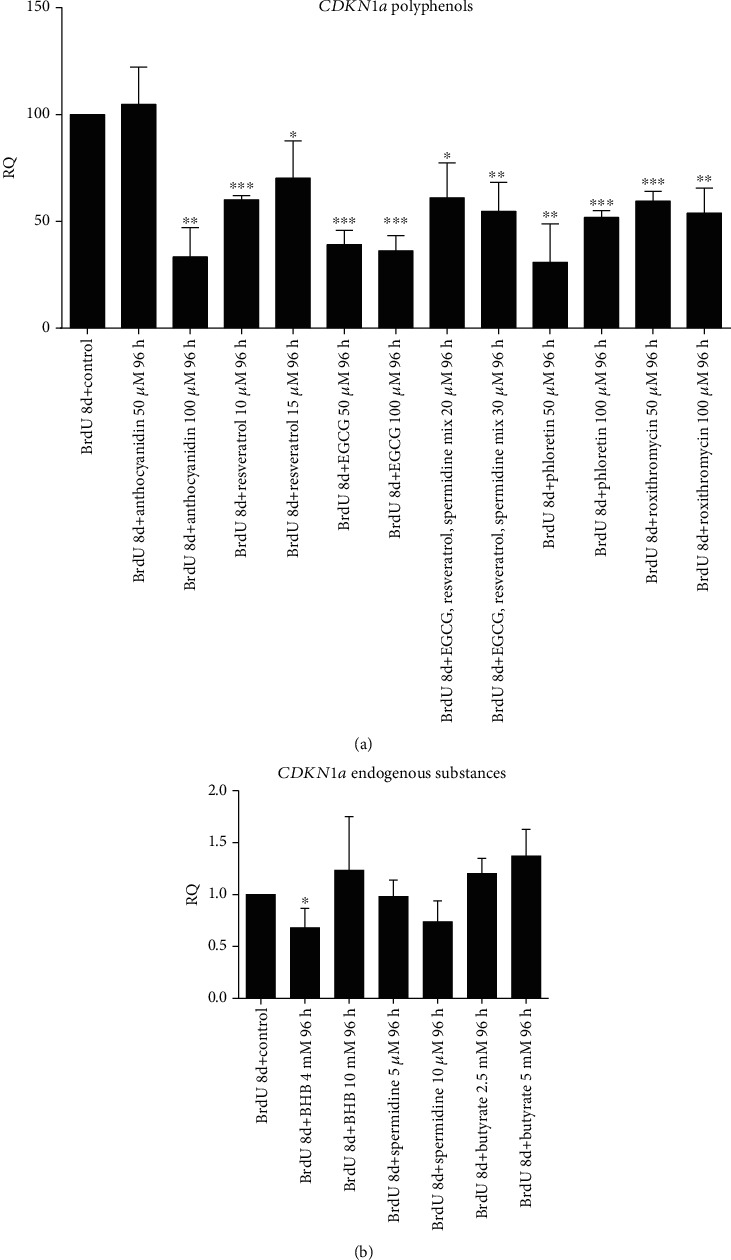
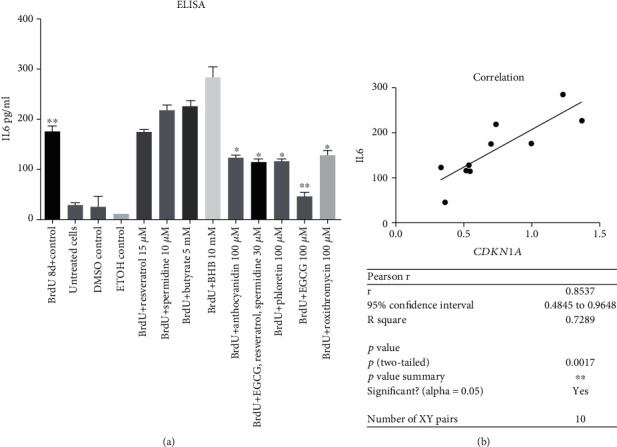
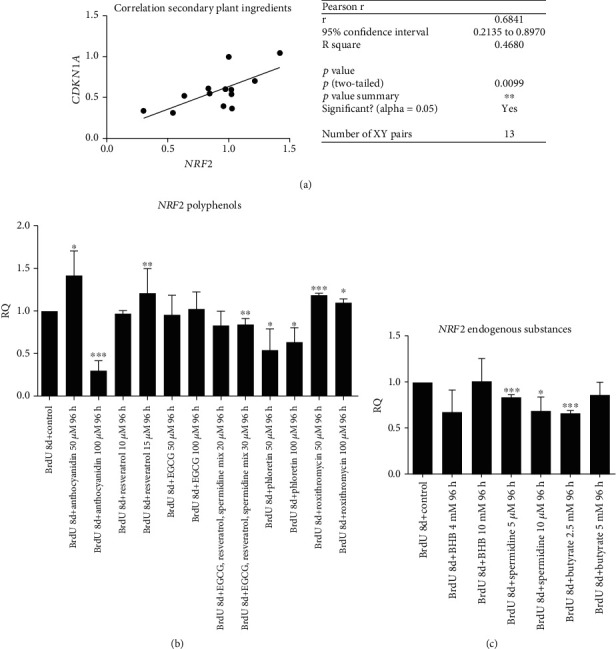
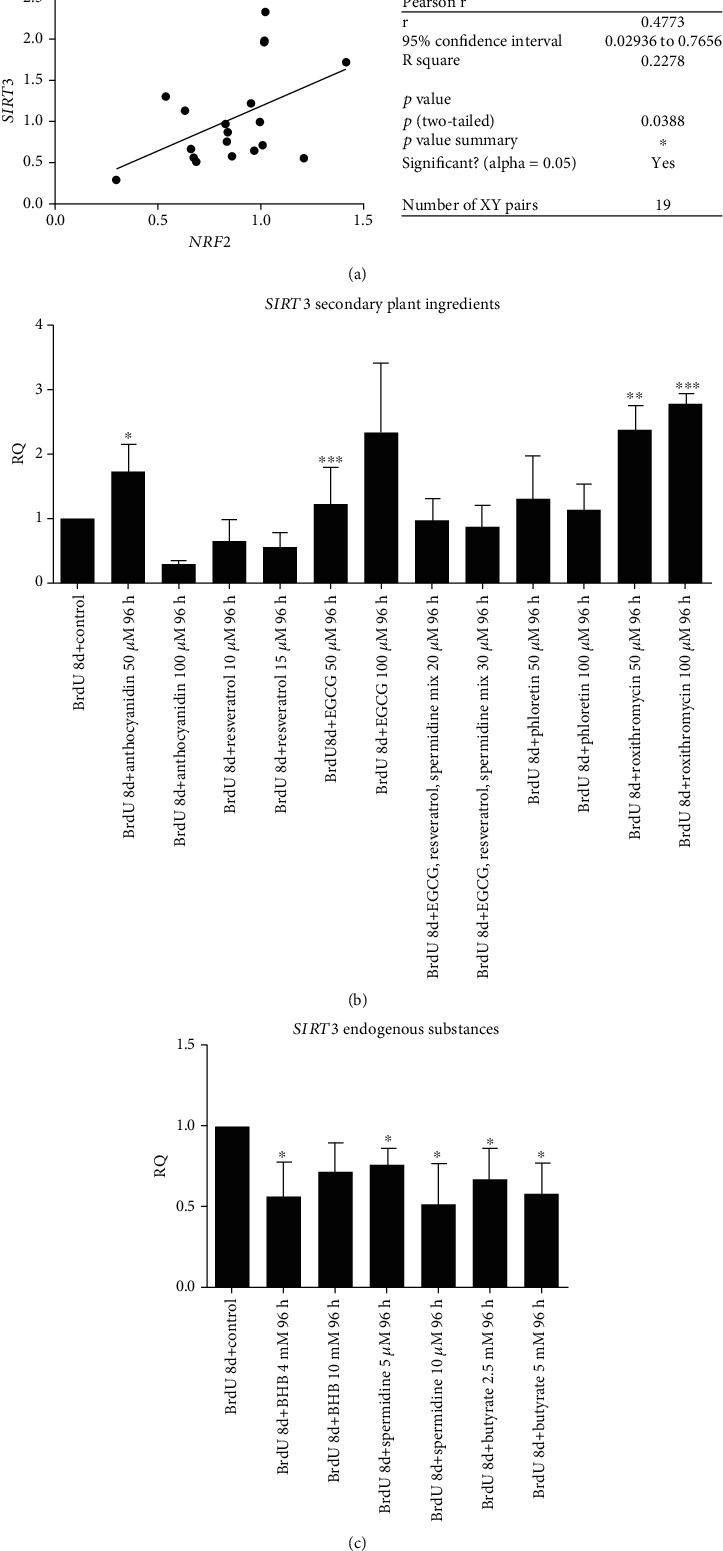
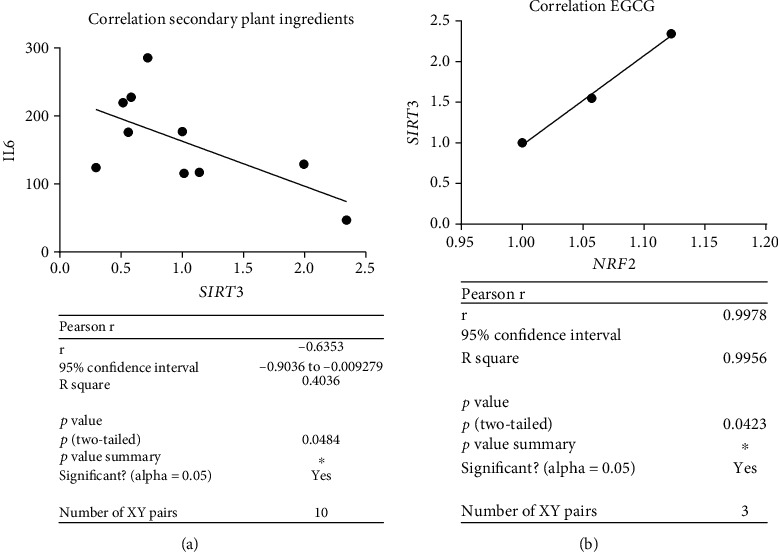
Results: CDKN1a increased significantly after BrdU incubation compared to untreated control (p < 0.01). All secondary plant ingredients used for treatment, but not anthocyanidin 50 μM, decrease CDKN1a expression (p < 0.05), whereas most endogenous substances did not attenuate CDKN1a. IL6 secretion positively correlated with CDKN1a (p < 0.01), whereas EGCG could diminish both, IL6 and CDKN1a with the strongest effect (p < 0.01). Although NRF2 positively correlated with SIRT3 activation (p < 0.05), only resveratrol (p < 0.01) and anthocyanidin (p < 0.05) could activate NRF2 significantly. Solely anthocyanidin 50 μM (p < 0.05) and 100 μM (p < 0.01) and EGCG 50 μM (p < 0.01) could increase SIRT3 expression. Activation of SIRT3 with EGCG correlated with lowered IL6 secretion significantly (p < 0.05) but not with anthocyanidin.
Conclusion: Accumulation of senescent cells in adipose tissue plays an important role in obesity and age-related diseases. SIRT3, located in the mitochondria, can regulate ROS via different pathways. Thus, targeting SIRT3 activating compounds such as EGCG may delay senescence of cells and senescence induced inflammatory processes.
Copyright © 2020 Stephanie Lilja et al.
Conflict of interest statement
The authors have no conflict of interest to declare.
Figures

Similar articles
-
Epigallocatechin gallate suppresses premature senescence of preadipocytes by inhibition of PI3K/Akt/mTOR pathway and induces senescent cell death by regulation of Bax/Bcl-2 pathway.Biogerontology. 2019 Apr;20(2):171-189. doi: 10.1007/s10522-018-9785-1. Epub 2018 Nov 19. Biogerontology. 2019. PMID: 30456590
-
Rapamycin inhibits the secretory phenotype of senescent cells by a Nrf2-independent mechanism.Aging Cell. 2017 Jun;16(3):564-574. doi: 10.1111/acel.12587. Epub 2017 Mar 31. Aging Cell. 2017. PMID: 28371119 Free PMC article.
-
Differential prooxidative effects of the green tea polyphenol, (-)-epigallocatechin-3-gallate, in normal and oral cancer cells are related to differences in sirtuin 3 signaling.Mol Nutr Food Res. 2015 Feb;59(2):203-11. doi: 10.1002/mnfr.201400485. Epub 2014 Nov 17. Mol Nutr Food Res. 2015. PMID: 25329972
-
Novel insights of dietary polyphenols and obesity.J Nutr Biochem. 2014 Jan;25(1):1-18. doi: 10.1016/j.jnutbio.2013.09.001. J Nutr Biochem. 2014. PMID: 24314860 Free PMC article. Review.
-
Cellular Senescence in Diabetes Mellitus: Distinct Senotherapeutic Strategies for Adipose Tissue and Pancreatic β Cells.Front Endocrinol (Lausanne). 2022 Mar 31;13:869414. doi: 10.3389/fendo.2022.869414. eCollection 2022. Front Endocrinol (Lausanne). 2022. PMID: 35432205 Free PMC article. Review.
Cited by
-
Nutraceutical Approaches of Autophagy and Neuroinflammation in Alzheimer's Disease: A Systematic Review.Molecules. 2020 Dec 18;25(24):6018. doi: 10.3390/molecules25246018. Molecules. 2020. PMID: 33353228 Free PMC article.
-
Crosstalk between adipogenesis and aging: role of polyphenols in combating adipogenic-associated aging.Immun Ageing. 2024 Nov 7;21(1):76. doi: 10.1186/s12979-024-00481-w. Immun Ageing. 2024. PMID: 39511615 Free PMC article. Review.
-
Targeting therapy-induced senescence as a novel strategy to combat chemotherapy-induced peripheral neuropathy.Support Care Cancer. 2024 Jan 5;32(1):85. doi: 10.1007/s00520-023-08287-0. Support Care Cancer. 2024. PMID: 38177894 Review.
-
Nutritional components as mitigators of cellular senescence in organismal aging: a comprehensive review.Food Sci Biotechnol. 2022 Jun 18;31(9):1089-1109. doi: 10.1007/s10068-022-01114-y. eCollection 2022 Aug. Food Sci Biotechnol. 2022. PMID: 35756719 Free PMC article. Review.
-
Profiling with senescence-associated secretory phenotype score identifies GDC-0879 as a small molecule sensitizing glioblastoma to anti-PD1.Cell Death Dis. 2025 Aug 9;16(1):602. doi: 10.1038/s41419-025-07915-3. Cell Death Dis. 2025. PMID: 40781221 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
