

Loss of Baiap2l2 destabilizes the transducing stereocilia of cochlear hair cells and leads to deafness
- PMID: 33151556
- PMCID: PMC7898316
- DOI: 10.1113/JP280670
Loss of Baiap2l2 destabilizes the transducing stereocilia of cochlear hair cells and leads to deafness
Abstract
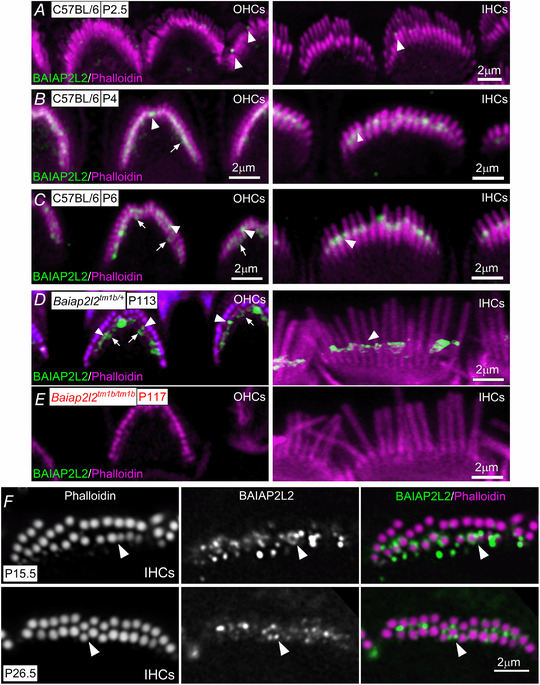
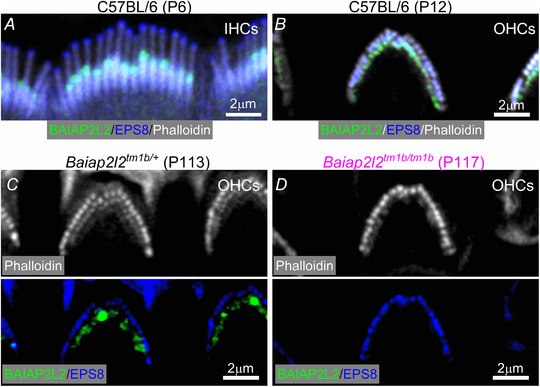
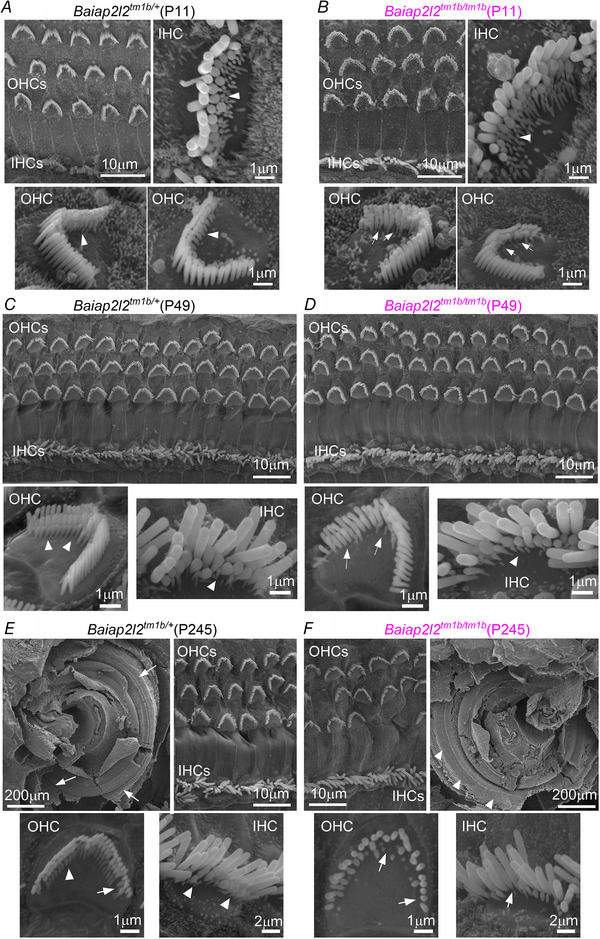
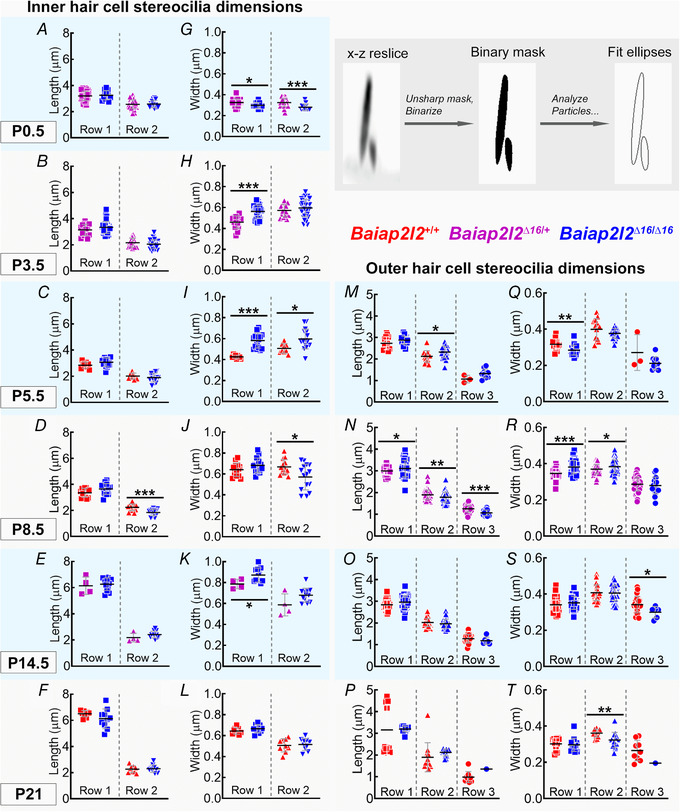
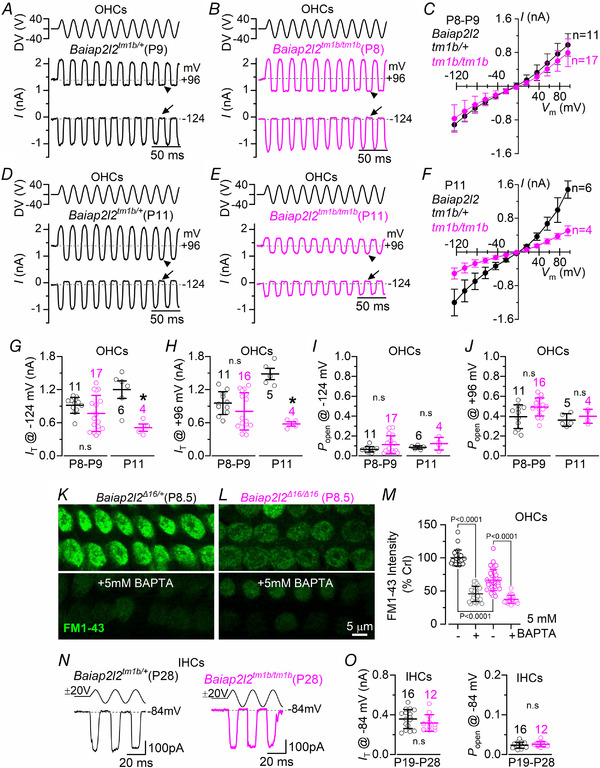
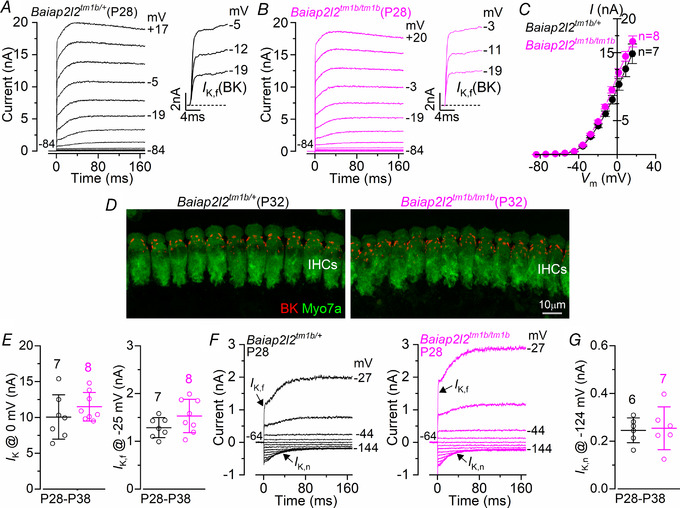
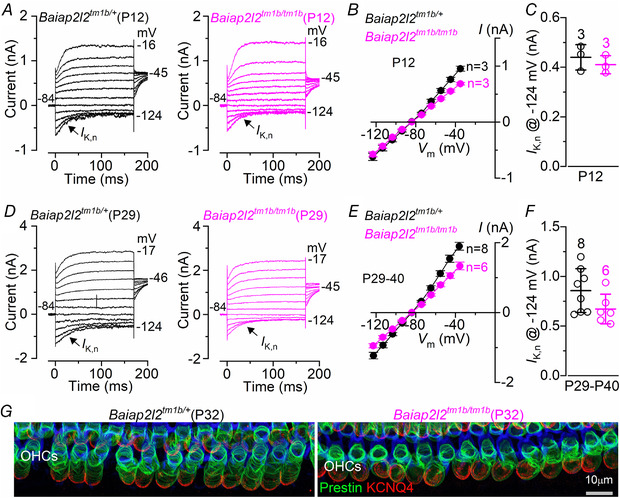
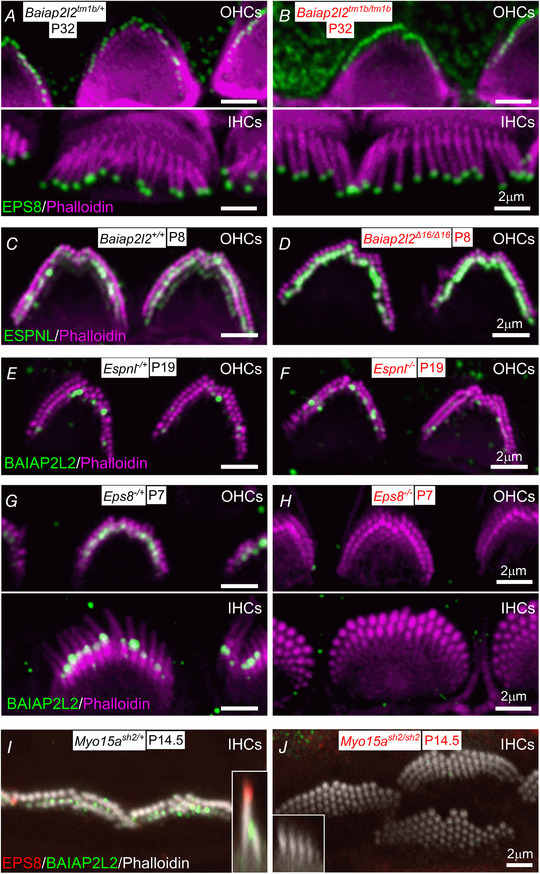
Key points: Mechanoelectrical transduction at auditory hair cells requires highly specialized stereociliary bundles that project from their apical surface, forming a characteristic graded 'staircase' structure. The morphogenesis and maintenance of these stereociliary bundles is a tightly regulated process requiring the involvement of several actin-binding proteins, many of which are still unidentified. We identify a new stereociliary protein, the I-BAR protein BAIAP2L2, which localizes to the tips of the shorter transducing stereocilia in both inner and outer hair cells (IHCs and OHCs). We find that Baiap2l2 deficient mice lose their second and third rows of stereocilia, their mechanoelectrical transducer current, and develop progressive hearing loss, becoming deaf by 8 months of age. We demonstrate that BAIAP2L2 localization to stereocilia tips is dependent on the motor protein MYO15A and its cargo EPS8. We propose that BAIAP2L2 is a new key protein required for the maintenance of the transducing stereocilia in mature cochlear hair cells.
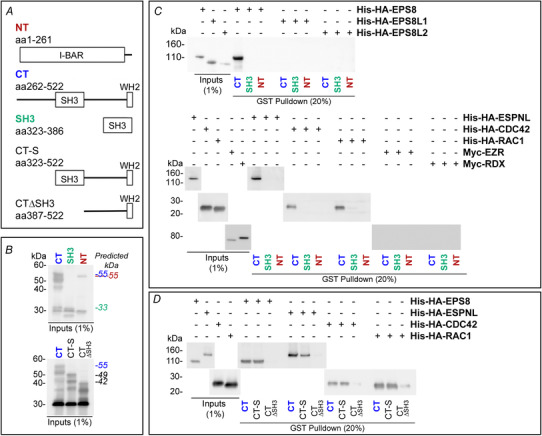
Abstract: The transduction of sound waves into electrical signals depends upon mechanosensitive stereociliary bundles that project from the apical surface of hair cells within the cochlea. The height and width of these actin-based stereocilia is tightly regulated throughout life to establish and maintain their characteristic staircase-like structure, which is essential for normal mechanoelectrical transduction. Here, we show that BAIAP2L2, a member of the I-BAR protein family, is a newly identified hair bundle protein that is localized to the tips of the shorter rows of transducing stereocilia in mouse cochlear hair cells. BAIAP2L2 was detected by immunohistochemistry from postnatal day 2.5 (P2.5) throughout adulthood. In Baiap2l2 deficient mice, outer hair cells (OHCs), but not inner hair cells (IHCs), began to lose their third row of stereocilia and showed a reduction in the size of the mechanoelectrical transducer current from just after P9. Over the following post-hearing weeks, the ordered staircase structure of the bundle progressively deteriorates, such that, by 8 months of age, both OHCs and IHCs of Baiap2l2 deficient mice have lost most of the second and third rows of stereocilia and become deaf. We also found that BAIAP2L2 interacts with other key stereociliary proteins involved in normal hair bundle morphogenesis, such as CDC42, RAC1, EPS8 and ESPNL. Furthermore, we show that BAIAP2L2 localization to the stereocilia tips depends on the motor protein MYO15A and its cargo EPS8. We propose that BAIAP2L2 is key to maintenance of the normal actin structure of the transducing stereocilia in mature mouse cochlear hair cells.
Keywords: actin; cochlear; development; hearing loss; mechanoelectrical transduction; mouse; stereocilia.
© 2020 The Authors. The Journal of Physiology published by John Wiley & Sons Ltd on behalf of The Physiological Society.
Figures
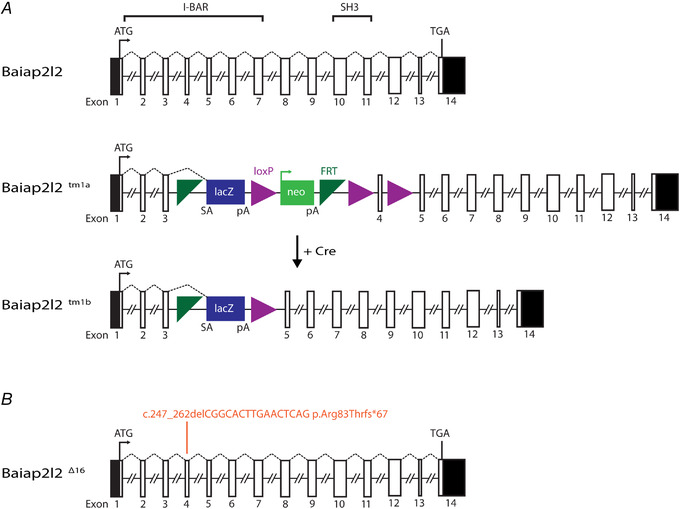


References
-
- Ahmed S, Goh WI & Bu W (2010). I‐BAR domains, IRSp53 and filopodium formation. Semin Cell Dev Biol 21, 350–356. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous