

PpCas9 from Pasteurella pneumotropica - a compact Type II-C Cas9 ortholog active in human cells
- PMID: 33152077
- PMCID: PMC7708072
- DOI: 10.1093/nar/gkaa998
PpCas9 from Pasteurella pneumotropica - a compact Type II-C Cas9 ortholog active in human cells
Abstract
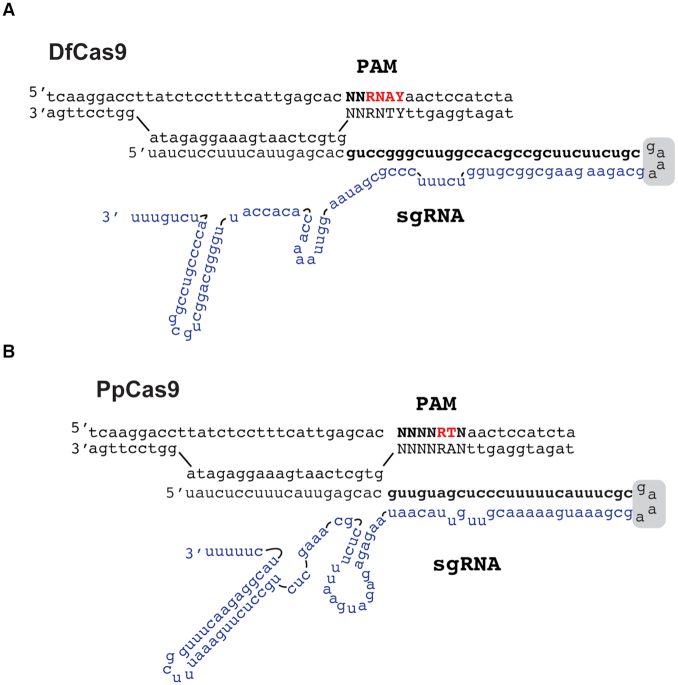
CRISPR-Cas defense systems opened up the field of genome editing due to the ease with which effector Cas nucleases can be programmed with guide RNAs to access desirable genomic sites. Type II-A SpCas9 from Streptococcus pyogenes was the first Cas9 nuclease used for genome editing and it remains the most popular enzyme of its class. Nevertheless, SpCas9 has some drawbacks including a relatively large size and restriction to targets flanked by an 'NGG' PAM sequence. The more compact Type II-C Cas9 orthologs can help to overcome the size limitation of SpCas9. Yet, only a few Type II-C nucleases were fully characterized to date. Here, we characterized two Cas9 II-C orthologs, DfCas9 from Defluviimonas sp.20V17 and PpCas9 from Pasteurella pneumotropica. Both DfCas9 and PpCas9 cleave DNA in vitro and have novel PAM requirements. Unlike DfCas9, the PpCas9 nuclease is active in human cells. This small nuclease requires an 'NNNNRTT' PAM orthogonal to that of SpCas9 and thus potentially can broaden the range of Cas9 applications in biomedicine and biotechnology.
© The Author(s) 2020. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures








Similar articles
-
Molecular basis for the PAM expansion and fidelity enhancement of an evolved Cas9 nuclease.PLoS Biol. 2019 Oct 11;17(10):e3000496. doi: 10.1371/journal.pbio.3000496. eCollection 2019 Oct. PLoS Biol. 2019. PMID: 31603896 Free PMC article.
-
Developing Heritable Mutations in Arabidopsis thaliana Using a Modified CRISPR/Cas9 Toolkit Comprising PAM-Altered Cas9 Variants and gRNAs.Plant Cell Physiol. 2019 Oct 1;60(10):2255-2262. doi: 10.1093/pcp/pcz118. Plant Cell Physiol. 2019. PMID: 31198958
-
DNA targeting by Clostridium cellulolyticum CRISPR-Cas9 Type II-C system.Nucleic Acids Res. 2020 Feb 28;48(4):2026-2034. doi: 10.1093/nar/gkz1225. Nucleic Acids Res. 2020. PMID: 31943070 Free PMC article.
-
[CRISPR/CAS9, the King of Genome Editing Tools].Mol Biol (Mosk). 2017 Jul-Aug;51(4):582-594. doi: 10.7868/S0026898417040036. Mol Biol (Mosk). 2017. PMID: 28900076 Review. Russian.
-
CRISPR-Cas systems: ushering in the new genome editing era.Bioengineered. 2018;9(1):214-221. doi: 10.1080/21655979.2018.1470720. Bioengineered. 2018. PMID: 29968520 Free PMC article. Review.
Cited by
-
Identification of the EH CRISPR-Cas9 system on a metagenome and its application to genome engineering.Microb Biotechnol. 2023 Jul;16(7):1505-1523. doi: 10.1111/1751-7915.14266. Epub 2023 Apr 25. Microb Biotechnol. 2023. PMID: 37097160 Free PMC article.
-
Full-Spectrum Targeted Mutagenesis in Plant and Animal Cells.Int J Mol Sci. 2021 Jan 16;22(2):857. doi: 10.3390/ijms22020857. Int J Mol Sci. 2021. PMID: 33467049 Free PMC article. Review.
-
Closely related type II-C Cas9 orthologs recognize diverse PAMs.Elife. 2022 Aug 12;11:e77825. doi: 10.7554/eLife.77825. Elife. 2022. PMID: 35959889 Free PMC article.
-
Identification of a novel type II-C Cas9 from the fish pathogen Flavobacterium psychrophilum.Front Microbiol. 2023 Jun 15;14:1181303. doi: 10.3389/fmicb.2023.1181303. eCollection 2023. Front Microbiol. 2023. PMID: 37396349 Free PMC article.
-
Massively parallel evaluation and computational prediction of the activities and specificities of 17 small Cas9s.Nat Methods. 2023 Jul;20(7):999-1009. doi: 10.1038/s41592-023-01875-2. Epub 2023 May 15. Nat Methods. 2023. PMID: 37188955
References
-
- Barrangou R., Fremaux C., Deveau H., Richards M., Boyaval P., Moineau S., Romero D.A., Horvath P.. CRISPR provides acquired resistance against viruses in prokaryotes. Science. 2007; 315:1709–1712. - PubMed
-
- Garneau J.E., Dupuis M.-È., Villion M., Romero D.A., Barrangou R., Boyaval P., Fremaux C., Horvath P., Magadán A.H., Moineau S.. The CRISPR/Cas bacterial immune system cleaves bacteriophage and plasmid DNA. Nature. 2010; 468:67–71. - PubMed
-
- Jansen R., van Embden J.D.A., Gaastra W., Schouls L.M.. Identification of genes that are associated with DNA repeats in prokaryotes. Mol. Microbiol. 2002; 43:1565–1575. - PubMed
-
- Pourcel C., Salvignol G., Vergnaud G.. CRISPR elements in Yersinia pestis acquire new repeats by preferential uptake of bacteriophage DNA, and provide additional tools for evolutionary studies. Microbiology (Reading). 2005; 151:653–663. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
