

The association of Epstein-Barr virus infection with CXCR3+ B-cell development in multiple sclerosis: impact of immunotherapies
- PMID: 33152118
- PMCID: PMC7984177
- DOI: 10.1002/eji.202048739
The association of Epstein-Barr virus infection with CXCR3+ B-cell development in multiple sclerosis: impact of immunotherapies
Abstract
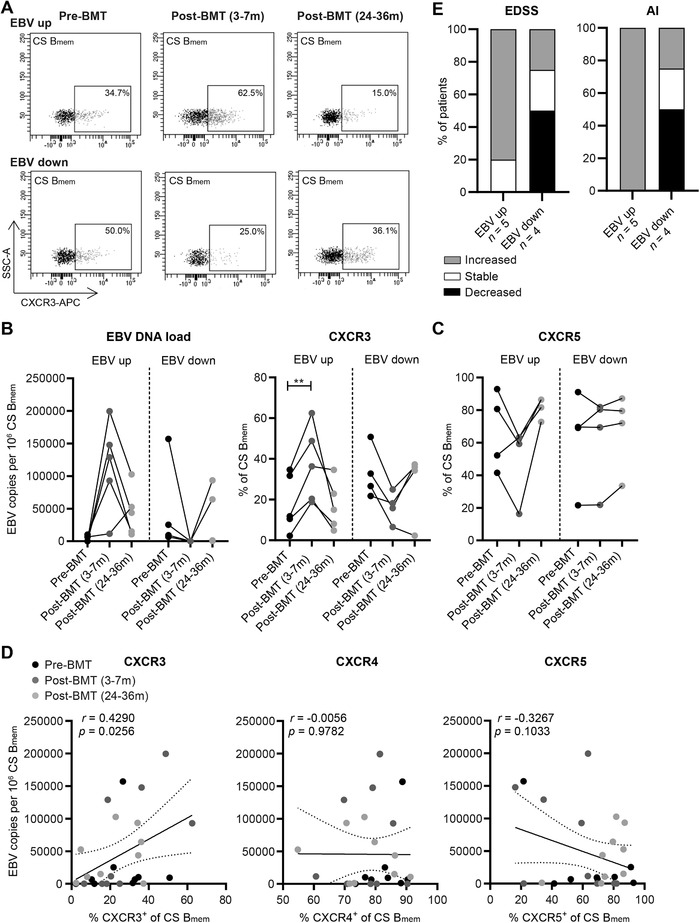
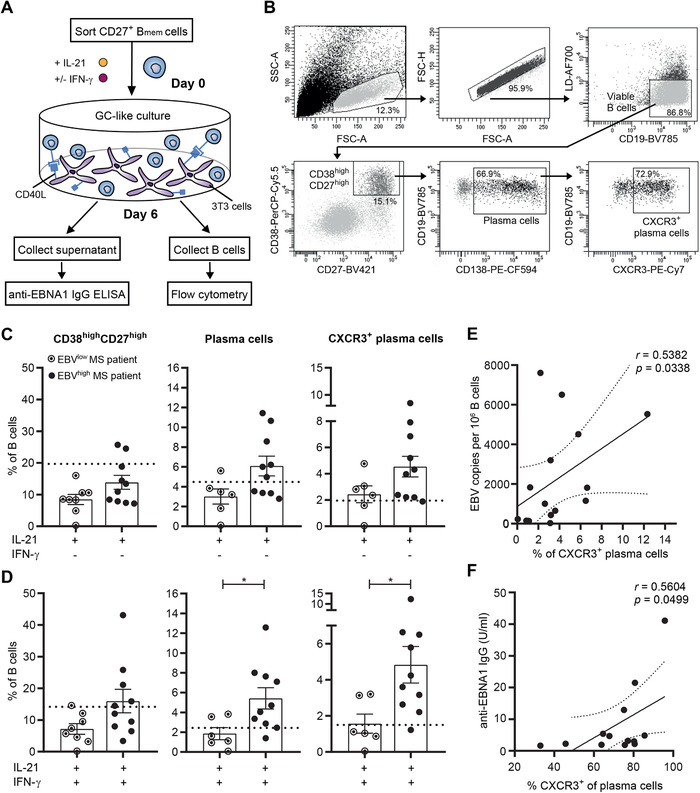
Epstein-Barr virus (EBV) infection of B cells is associated with increased multiple sclerosis (MS) susceptibility. Recently, we found that CXCR3-expressing B cells preferentially infiltrate the CNS of MS patients. In chronic virus-infected mice, these types of B cells are sustained and show increased antiviral responsiveness. How EBV persistence in B cells influences their development remains unclear. First, we analyzed ex vivo B-cell subsets from MS patients who received autologous bone marrow transplantation (n = 9), which is often accompanied by EBV reactivation. The frequencies of nonclass-switched and class-switched memory B cells were reduced at 3-7 months, while only class-switched B cells returned back to baseline at 24-36 months posttransplantation. At these time points, EBV DNA load positively correlated to the frequency of CXCR3+ , and not CXCR4+ or CXCR5+ , class-switched B cells. Second, for CXCR3+ memory B cells trapped within the blood of MS patients treated with natalizumab (anti-VLA-4 antibody n = 15), latent EBV infection corresponded to enhanced in vitro formation of anti-EBNA1 IgG-secreting plasma cells under GC-like conditions. These findings imply that EBV persistence in B cells potentiates brain-homing and antibody-producing CXCR3+ subsets in MS.
Keywords: EBV; memory B cells; multiple sclerosis; natalizumab; plasma cells.
© 2020 The Authors. European Journal of Immunology published by Wiley-VCH GmbH.
Conflict of interest statement
J.S. received speaker/consultancy fee from Biogen, Merck, Novartis, and Sanofi‐Genzyme. All other authors have no commercial or financial conflicts of interest.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
