

Structural basis of antagonizing the vitamin K catalytic cycle for anticoagulation
- PMID: 33154105
- PMCID: PMC7946407
- DOI: 10.1126/science.abc5667
Structural basis of antagonizing the vitamin K catalytic cycle for anticoagulation
Abstract
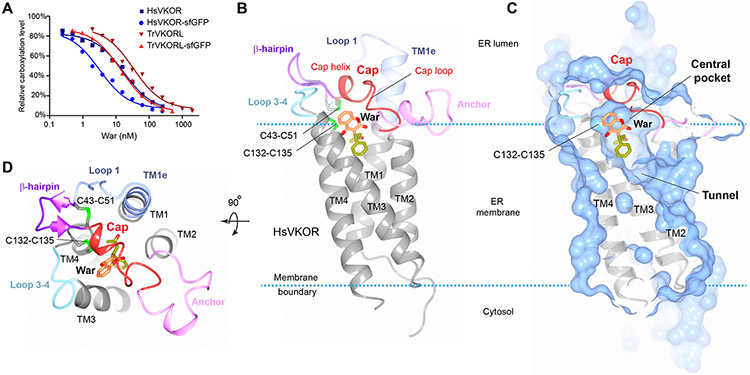
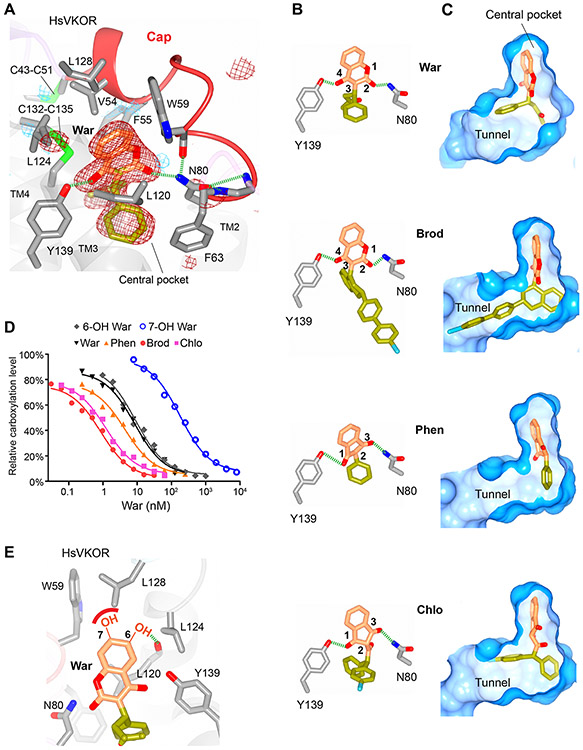
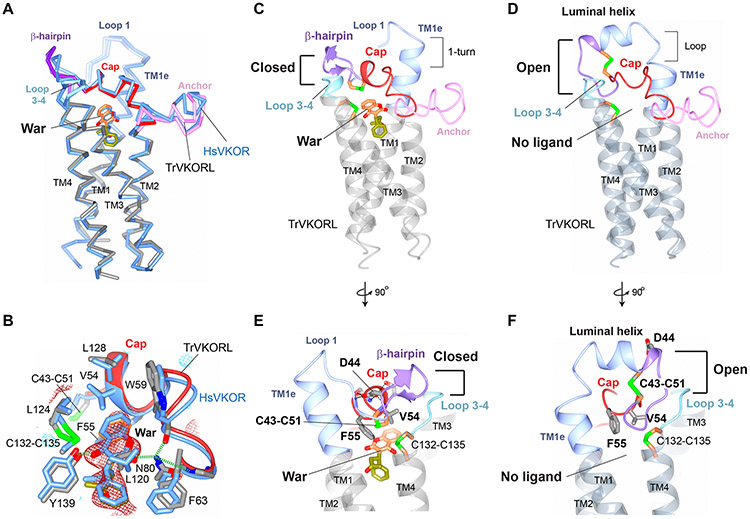
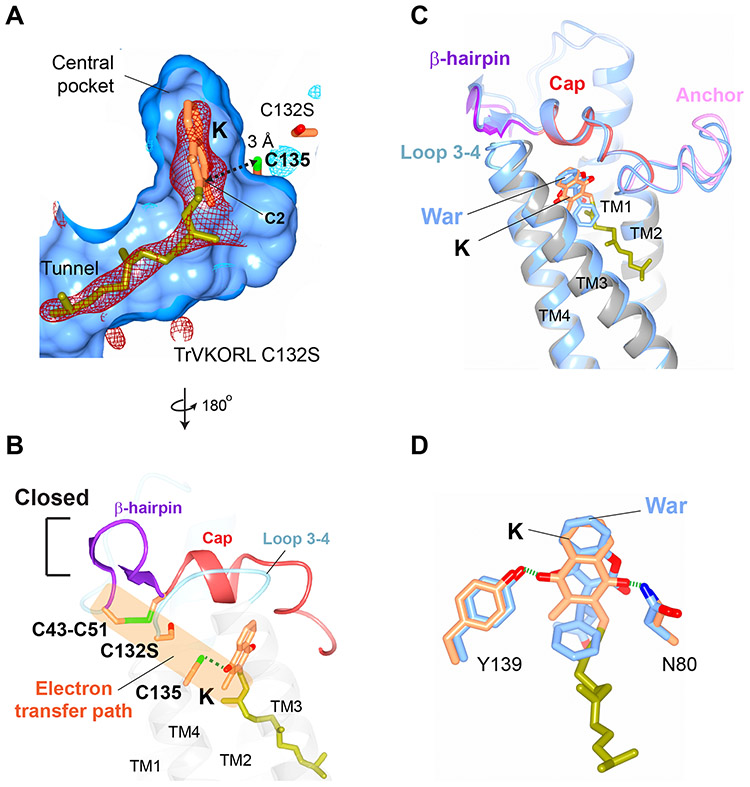
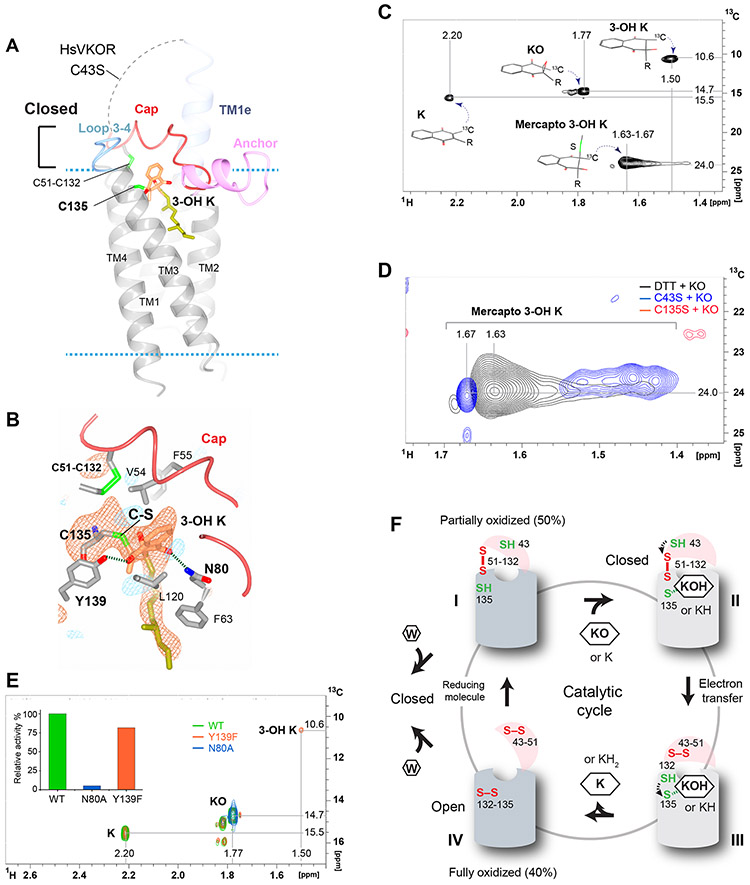
Vitamin K antagonists are widely used anticoagulants that target vitamin K epoxide reductases (VKOR), a family of integral membrane enzymes. To elucidate their catalytic cycle and inhibitory mechanism, we report 11 x-ray crystal structures of human VKOR and pufferfish VKOR-like, with substrates and antagonists in different redox states. Substrates entering the active site in a partially oxidized state form cysteine adducts that induce an open-to-closed conformational change, triggering reduction. Binding and catalysis are facilitated by hydrogen-bonding interactions in a hydrophobic pocket. The antagonists bind specifically to the same hydrogen-bonding residues and induce a similar closed conformation. Thus, vitamin K antagonists act through mimicking the key interactions and conformational changes required for the VKOR catalytic cycle.
Copyright © 2021, American Association for the Advancement of Science.
Figures
Similar articles
-
Structural and cellular basis of vitamin K antagonism.J Thromb Haemost. 2022 Sep;20(9):1971-1983. doi: 10.1111/jth.15800. Epub 2022 Jul 12. J Thromb Haemost. 2022. PMID: 35748323 Review.
-
Structures of an intramembrane vitamin K epoxide reductase homolog reveal control mechanisms for electron transfer.Nat Commun. 2014;5:3110. doi: 10.1038/ncomms4110. Nat Commun. 2014. PMID: 24477003 Free PMC article.
-
Structural features determining the vitamin K epoxide reduction activity in the VKOR family of membrane oxidoreductases.FEBS J. 2022 Aug;289(15):4564-4579. doi: 10.1111/febs.16386. Epub 2022 Feb 10. FEBS J. 2022. PMID: 35113495 Free PMC article.
-
Warfarin and vitamin K compete for binding to Phe55 in human VKOR.Nat Struct Mol Biol. 2017 Jan;24(1):77-85. doi: 10.1038/nsmb.3338. Epub 2016 Dec 12. Nat Struct Mol Biol. 2017. PMID: 27941861
-
Structural and functional insights into enzymes of the vitamin K cycle.J Thromb Haemost. 2016 Feb;14(2):236-47. doi: 10.1111/jth.13217. Epub 2016 Jan 29. J Thromb Haemost. 2016. PMID: 26663892 Free PMC article. Review.
Cited by
-
A physiologically-based pharmacokinetic/pharmacodynamic modeling approach for drug-drug-gene interaction evaluation of S-warfarin with fluconazole.CPT Pharmacometrics Syst Pharmacol. 2024 May;13(5):853-869. doi: 10.1002/psp4.13123. Epub 2024 Mar 15. CPT Pharmacometrics Syst Pharmacol. 2024. PMID: 38487942 Free PMC article.
-
Termini restraining of small membrane proteins enables structure determination at near-atomic resolution.Sci Adv. 2020 Dec 18;6(51):eabe3717. doi: 10.1126/sciadv.abe3717. Print 2020 Dec. Sci Adv. 2020. PMID: 33355146 Free PMC article.
-
Yeast as a tool for membrane protein production and structure determination.FEMS Yeast Res. 2022 Oct 20;22(1):foac047. doi: 10.1093/femsyr/foac047. FEMS Yeast Res. 2022. PMID: 36175165 Free PMC article.
-
New polymorphisms of Vkork1 gene related to anticoagulant resistance of rats and mice in Italy.Pest Manag Sci. 2025 Jun;81(6):2869-2880. doi: 10.1002/ps.8652. Epub 2025 Jan 15. Pest Manag Sci. 2025. PMID: 39815713 Free PMC article.
-
In Silico and In Vitro Studies of Potential Novel Vitamin K Epoxide Reductase (VKOR) Inhibitors Suggest an Updated Structure-Activity Relationship.ACS Omega. 2025 Jun 13;10(25):26694-26704. doi: 10.1021/acsomega.5c01116. eCollection 2025 Jul 1. ACS Omega. 2025. PMID: 40621007 Free PMC article.
References
-
- Goy J, Crowther M, Approaches to diagnosing and managing anticoagulant-related bleeding. Seminars in Thrombosis and Hemostasis 38, 702–710 (2012). - PubMed
-
- Schulman S, Beyth RJ, Kearon C, Levine MN, Hemorrhagic complications of anticoagulant and thrombolytic treatment: American College of Chest Physicians evidence-based clinical practice guidelines (8th edition). (2008), vol. 133. - PubMed
-
- Budnitz DS, Lovegrove MC, Shehab N, Richards CL, Emergency hospitalizations for adverse drug events in older Americans. The New England journal of medicine 365, 2002–2012 (2011). - PubMed
-
- Stafford DW, The vitamin K cycle. J Thromb Haemost 3, 1873–1878 (2005). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
