

Alzheimer's amyloid β heterogeneous species differentially affect brain endothelial cell viability, blood-brain barrier integrity, and angiogenesis
- PMID: 33155752
- PMCID: PMC7681048
- DOI: 10.1111/acel.13258
Alzheimer's amyloid β heterogeneous species differentially affect brain endothelial cell viability, blood-brain barrier integrity, and angiogenesis
Abstract
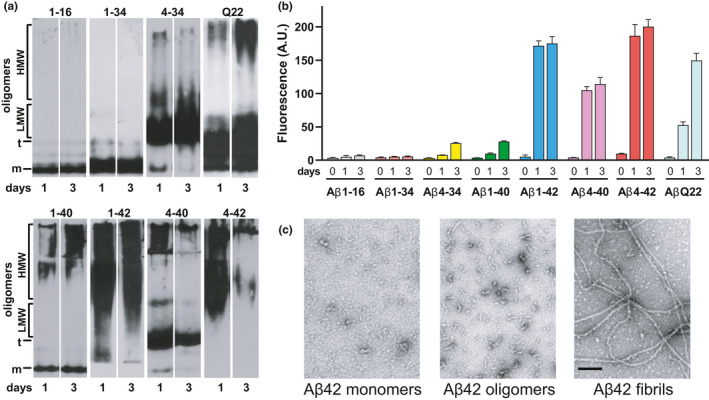
Impaired clearance in the Alzheimer's Disease (AD) brain is key in the formation of Aβ parenchymal plaques and cerebrovascular deposits known as cerebral amyloid angiopathy (CAA), present in >80% of AD patients and ~50% of non-AD elderly subjects. Aβ deposits are highly heterogeneous, containing multiple fragments mostly derived from catabolism of Aβ40/Aβ42, which exhibit dissimilar aggregation properties. Remarkably, the role of these physiologically relevant Aβ species in cerebrovascular injury and their impact in vascular pathology is unknown. We sought to understand how heterogeneous Aβ species affect cerebral endothelial health and assess whether their diverse effects are associated with the peptides aggregation propensities. We analyzed cerebral microvascular endothelial cell (CMEC) viability, blood-brain barrier (BBB) permeability, and angiogenesis, all relevant aspects of brain microvascular dysfunction. We found that Aβ peptides and fragments exerted differential effects on cerebrovascular pathology. Peptides forming mostly oligomeric structures induced CMEC apoptosis, whereas fibrillar aggregates increased BBB permeability without apoptotic effects. Interestingly, all Aβ species tested inhibited angiogenesis in vitro. These data link the biological effects of the heterogeneous Aβ peptides to their primary structure and aggregation, strongly suggesting that the composition of amyloid deposits influences clinical aspects of the AD vascular pathology. As the presence of predominant oligomeric structures in proximity of the vessel walls may lead to CMEC death and induction of microhemorrhages, fibrillar amyloid is likely responsible for increased BBB permeability and associated neurovascular dysfunction. These results have the potential to unveil more specific therapeutic targets and clarify the multifactorial nature of AD.
Keywords: Alzheimer's disease; amyloid β; angiogenesis; blood-brain barrier; cerebral amyloid angiopathy.
© 2020 The Authors. Aging Cell published by the Anatomical Society and John Wiley & Sons Ltd.
Conflict of interest statement
None.
Figures





Similar articles
-
Impaired Amyloid Beta Clearance and Brain Microvascular Dysfunction are Present in the Tg-SwDI Mouse Model of Alzheimer's Disease.Neuroscience. 2020 Aug 1;440:48-55. doi: 10.1016/j.neuroscience.2020.05.024. Epub 2020 May 23. Neuroscience. 2020. PMID: 32450297
-
Traffic jam at the blood-brain barrier promotes greater accumulation of Alzheimer's disease amyloid-β proteins in the cerebral vasculature.Mol Pharm. 2013 May 6;10(5):1557-65. doi: 10.1021/mp300352c. Epub 2013 Mar 5. Mol Pharm. 2013. PMID: 23249146 Free PMC article.
-
Homocysteine potentiates amyloid -induced death receptor 4- and 5-mediated cerebral endothelial cell apoptosis, blood brain barrier dysfunction and angiogenic impairment.Aging Cell. 2024 May;23(5):e14106. doi: 10.1111/acel.14106. Epub 2024 Feb 15. Aging Cell. 2024. PMID: 38358083 Free PMC article.
-
Endothelial Mitochondrial Dysfunction in Cerebral Amyloid Angiopathy and Alzheimer's Disease.J Alzheimers Dis. 2019;72(4):1019-1039. doi: 10.3233/JAD-190357. J Alzheimers Dis. 2019. PMID: 31306129 Free PMC article. Review.
-
Neurovascular mechanisms and blood-brain barrier disorder in Alzheimer's disease.Acta Neuropathol. 2009 Jul;118(1):103-13. doi: 10.1007/s00401-009-0522-3. Epub 2009 Mar 25. Acta Neuropathol. 2009. PMID: 19319544 Free PMC article. Review.
Cited by
-
The Effects of Amyloid-β on Metabolomic Profiles of Cardiomyocytes and Coronary Endothelial Cells.J Alzheimers Dis. 2023;93(1):307-319. doi: 10.3233/JAD-221199. J Alzheimers Dis. 2023. PMID: 36970904 Free PMC article.
-
Comparison of Cerebrospinal Fluid Amyloidogenic Nanoplaques With Core Biomarkers of Alzheimer's Disease.Front Aging Neurosci. 2021 Jan 8;12:608628. doi: 10.3389/fnagi.2020.608628. eCollection 2020. Front Aging Neurosci. 2021. PMID: 33488383 Free PMC article.
-
Lipidopathy disrupts peripheral and central amyloid clearance in Alzheimer's disease: Where are our knowledge.IBRO Neurosci Rep. 2025 Jan 9;18:191-199. doi: 10.1016/j.ibneur.2025.01.004. eCollection 2025 Jun. IBRO Neurosci Rep. 2025. PMID: 39906286 Free PMC article. Review.
-
Fibrillar tau alters cerebral endothelial cell metabolism, vascular inflammatory activation, and barrier function in vitro and in vivo.Alzheimers Dement. 2025 Mar;21(3):e70077. doi: 10.1002/alz.70077. Alzheimers Dement. 2025. PMID: 40110691 Free PMC article.
-
Electroacupuncture ameliorates cerebrovascular impairment in Alzheimer's disease mice via melatonin signaling.CNS Neurosci Ther. 2023 Mar;29(3):917-931. doi: 10.1111/cns.14027. Epub 2022 Nov 15. CNS Neurosci Ther. 2023. PMID: 36382345 Free PMC article.
References
-
- Bakker, E. N. , Bacskai, B. J. , Arbel‐Ornath, M. , Aldea, R. , Bedussi, B. , Morris, A. W. , Weller, R. O. , & Carare, R. O. (2016). Lymphatic clearance of the brain: Perivascular, paravascular and significance for neurodegenerative diseases. Cellular and Molecular Neurobiology, 36(2), 181–194. 10.1007/s10571-015-0273-8 - DOI - PMC - PubMed
-
- Benilova, I. , Karran, E. , & De Strooper, B. (2012). The toxic Abeta oligomer and Alzheimer's disease: An emperor in need of clothes. Nature Neuroscience, 15, 349–357. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
