

Coordinate Regulation of Ribosome and tRNA Biogenesis Controls Hypoxic Injury and Translation
- PMID: 33157031
- PMCID: PMC7855744
- DOI: 10.1016/j.cub.2020.10.001
Coordinate Regulation of Ribosome and tRNA Biogenesis Controls Hypoxic Injury and Translation
Erratum in
-
Coordinate Regulation of Ribosome and tRNA Biogenesis Controls Hypoxic Injury and Translation.Curr Biol. 2022 Dec 19;32(24):5433. doi: 10.1016/j.cub.2022.11.053. Curr Biol. 2022. PMID: 36538878 Free PMC article. No abstract available.
Abstract
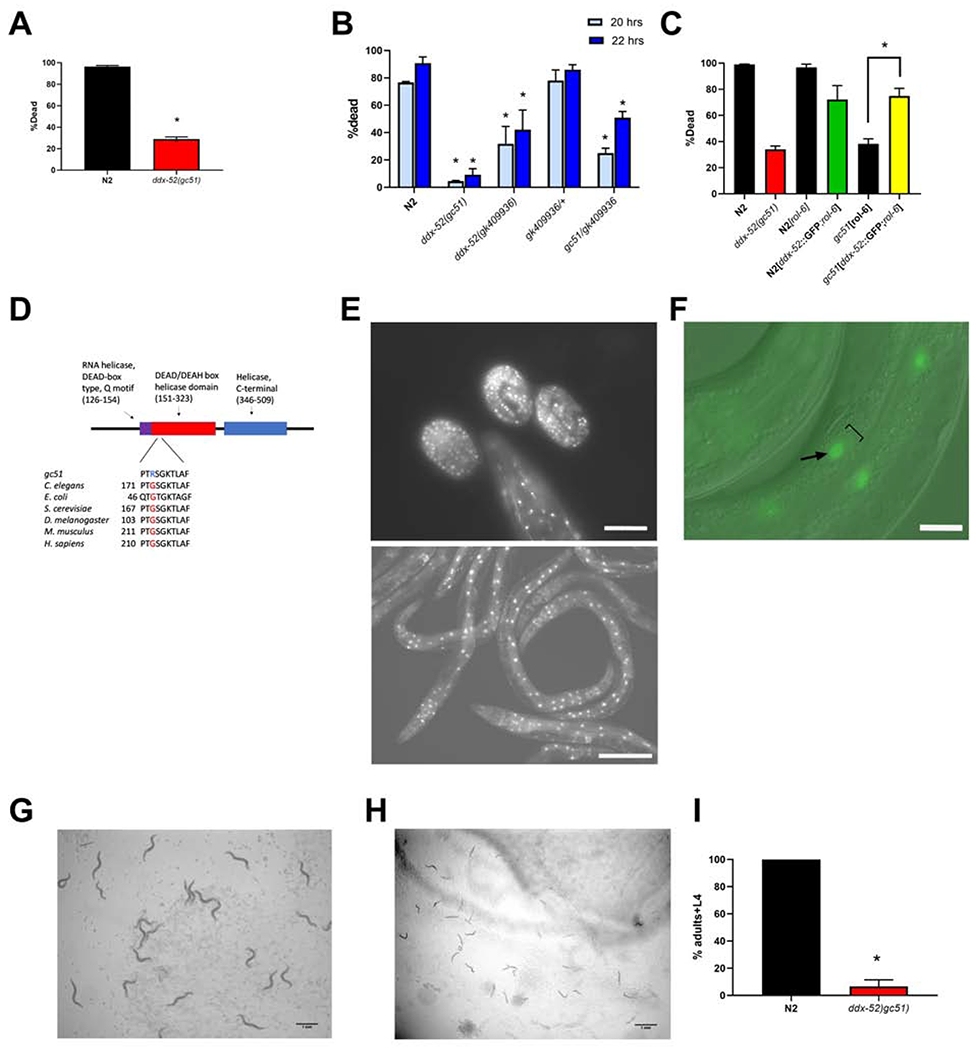
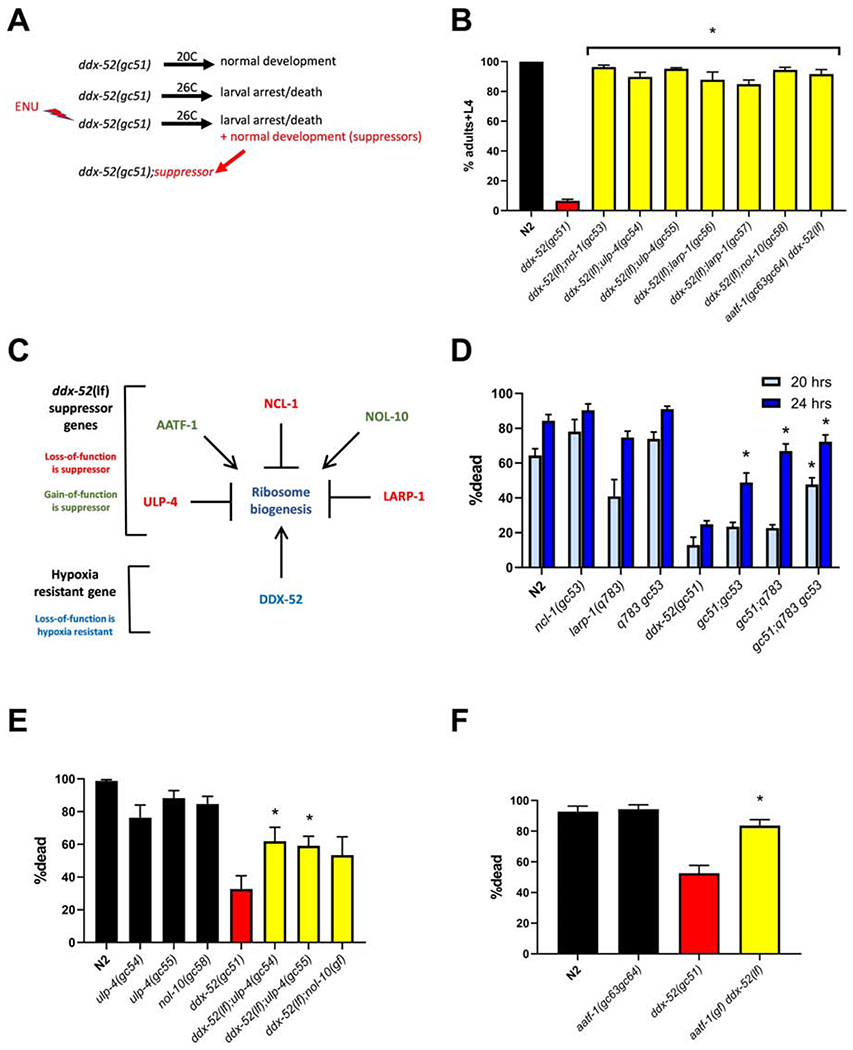
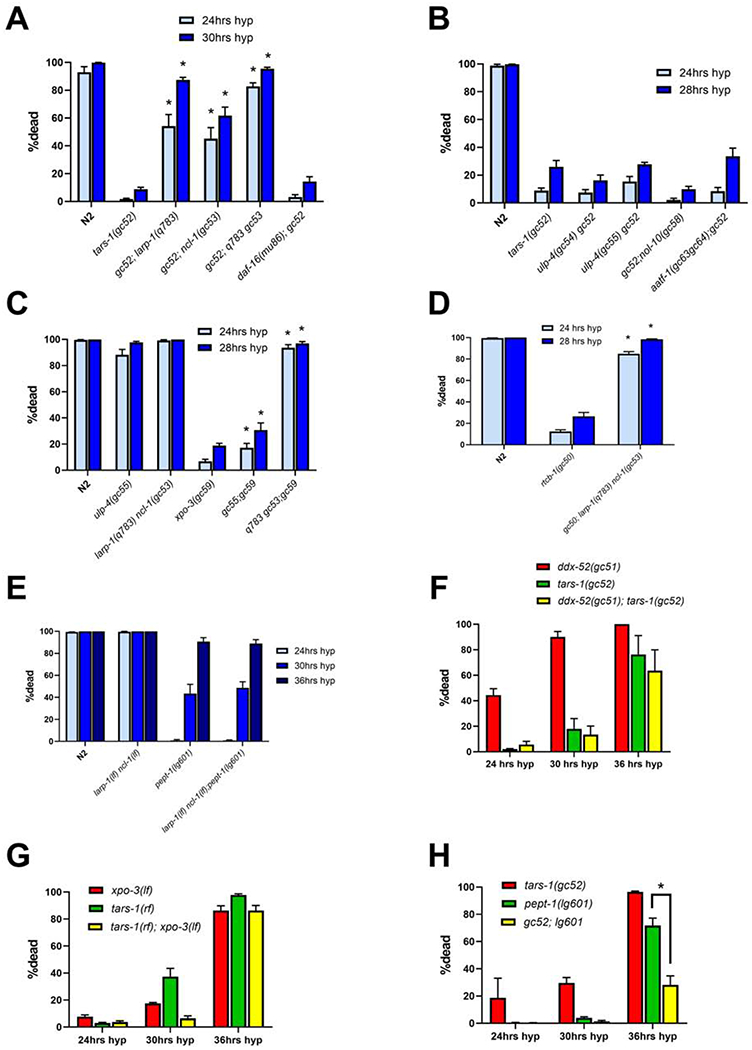
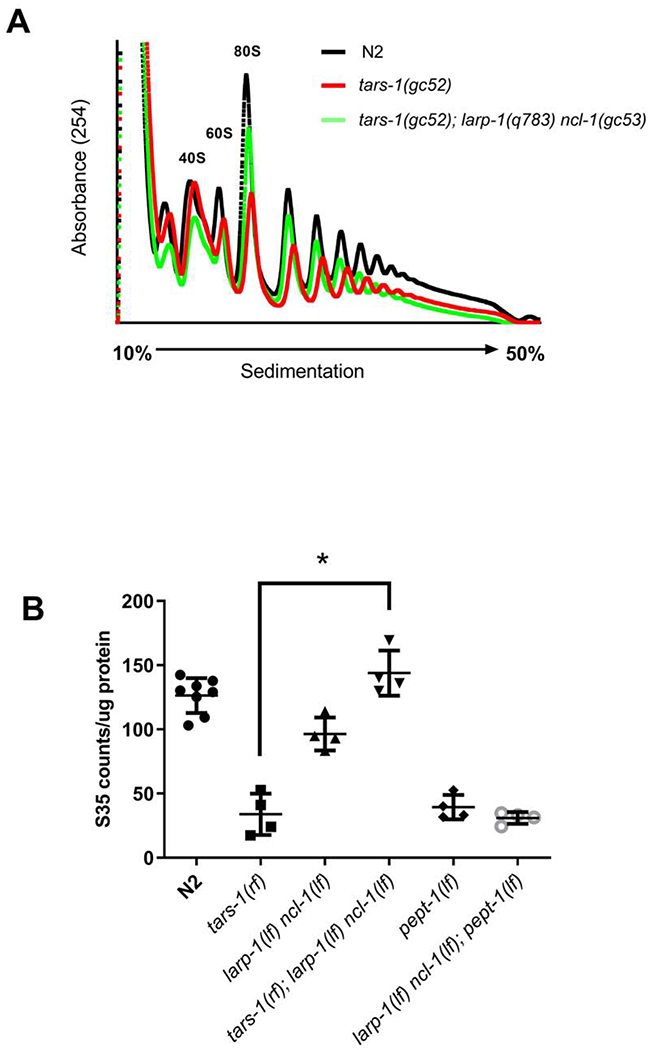
The translation machinery is composed of a myriad of proteins and RNAs whose levels must be coordinated to efficiently produce proteins without wasting energy or substrate. However, protein synthesis is clearly not always perfectly tuned to its environment, as disruption of translation machinery components can lengthen lifespan and stress survival. While much has been learned from bacteria and yeast about translational regulation, much less is known in metazoans. In a screen for mutations protecting C. elegans from hypoxic stress, we isolated multiple genes impacting protein synthesis: a ribosomal RNA helicase gene, tRNA biosynthesis genes, and a gene controlling amino acid availability. To define better the mechanisms by which these genes impact protein synthesis, we performed a second screen for suppressors of the conditional developmental arrest phenotype of the RNA helicase mutant and identified genes involved in ribosome biogenesis. Surprisingly, these suppressor mutations restored normal hypoxic sensitivity and protein synthesis to the tRNA biogenesis mutants, but not to the mutant reducing amino acid uptake. Proteomic analysis demonstrated that reduced tRNA biosynthetic activity produces a selective homeostatic reduction in ribosomal subunits, thereby offering a mechanism for the suppression results. Our study uncovers an unrecognized higher-order-translation regulatory mechanism in a metazoan whereby ribosome biogenesis genes communicate with genes controlling tRNA abundance matching the global rate of protein synthesis with available resources.
Keywords: aminoacyl tRNA synthetase; hypoxic cell death; protein synthesis; proteomics; ribosomal; ribosomal RNA helicase; tRNA ligase.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
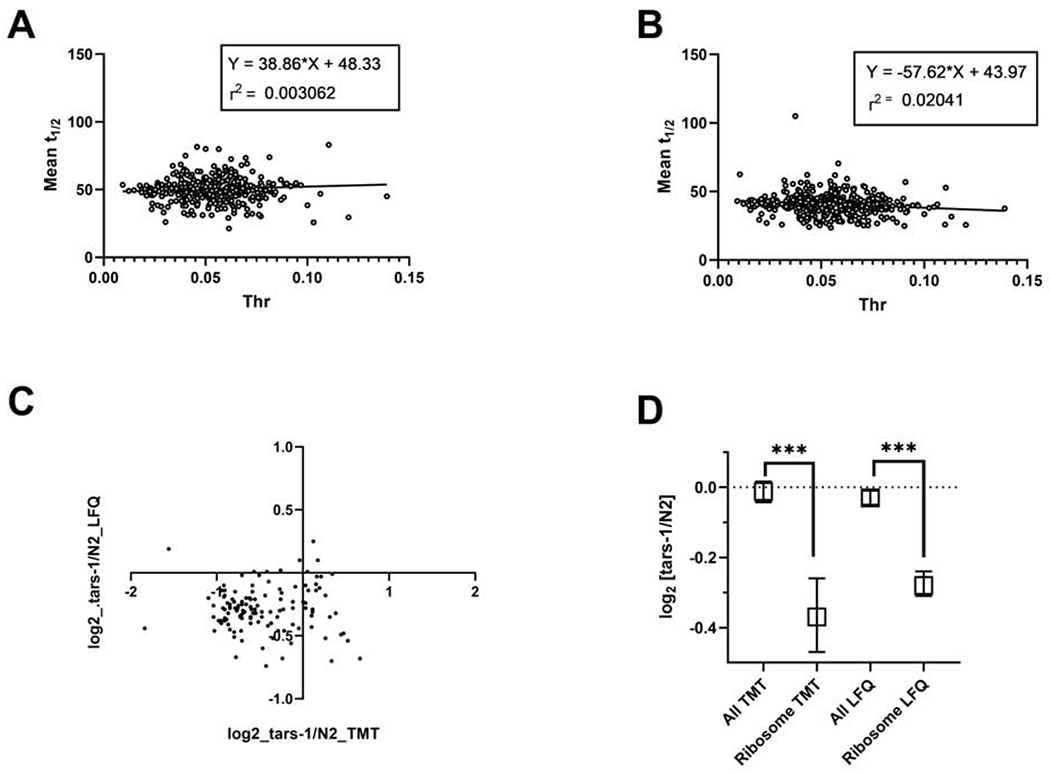
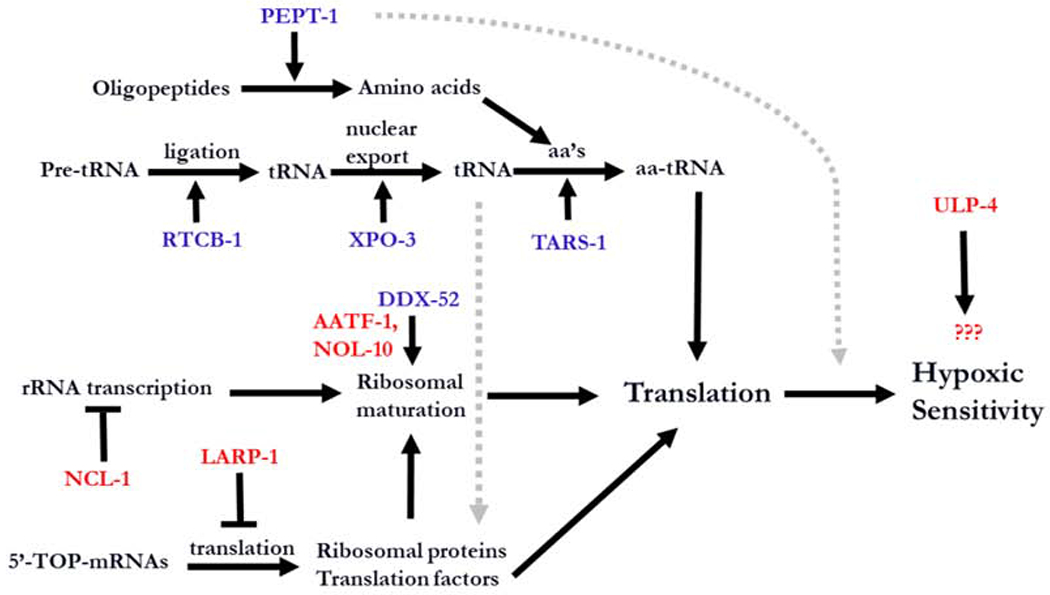
Comment in
-
Hypoxia: Uncharged tRNA to the Rescue!Curr Biol. 2021 Jan 11;31(1):R25-R27. doi: 10.1016/j.cub.2020.10.067. Curr Biol. 2021. PMID: 33434482
References
-
- Spriggs KA, Bushell M, and Willis AE (2010). Translational regulation of gene expression during conditions of cell stress. Mol Cell 40, 228–237. - PubMed
-
- Romero-Ruiz A, Bautista L, Navarro V, Heras-Garvin A, March-Diaz R, Castellano A, Gomez-Diaz R, Castro MJ, Berra E, Lopez-Barneo J, et al. (2012). Prolyl hydroxylase-dependent modulation of eukaryotic elongation factor 2 activity and protein translation under acute hypoxia. J Biol Chem 287, 9651–9658. - PMC - PubMed
-
- Robichaud N, and Sonenberg N (2017). Translational control and the cancer cell response to stress. Curr Opin Cell Biol 45, 102–109. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
