

The nucleocytosolic O-fucosyltransferase SPINDLY affects protein expression and virulence in Toxoplasma gondii
- PMID: 33158988
- PMCID: PMC7949088
- DOI: 10.1074/jbc.RA120.015883
The nucleocytosolic O-fucosyltransferase SPINDLY affects protein expression and virulence in Toxoplasma gondii
Abstract
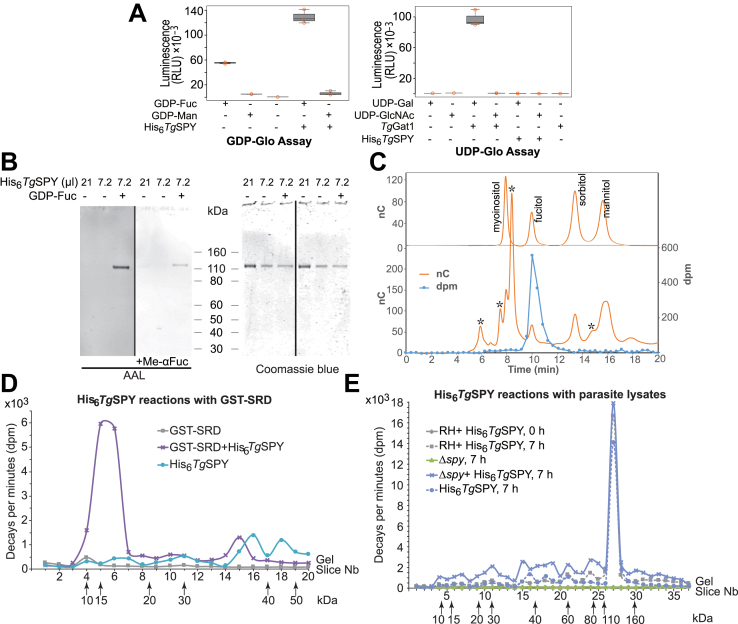
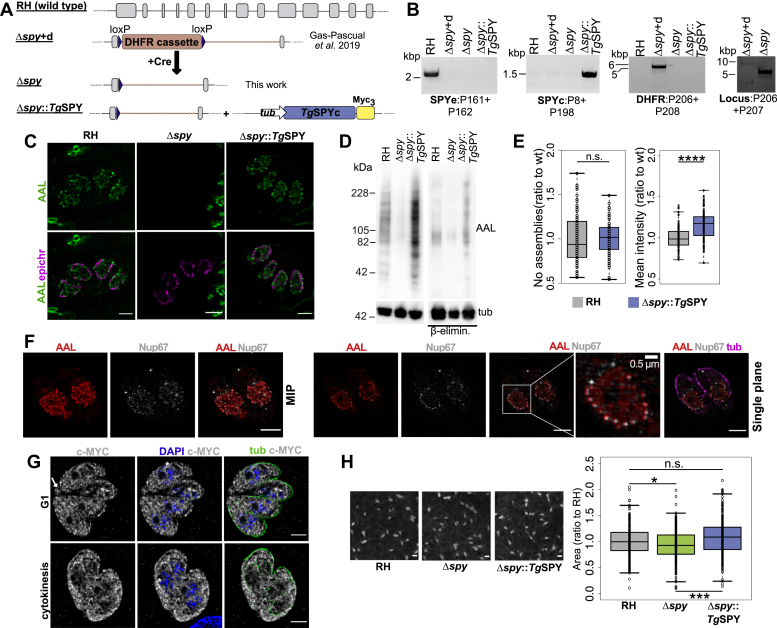
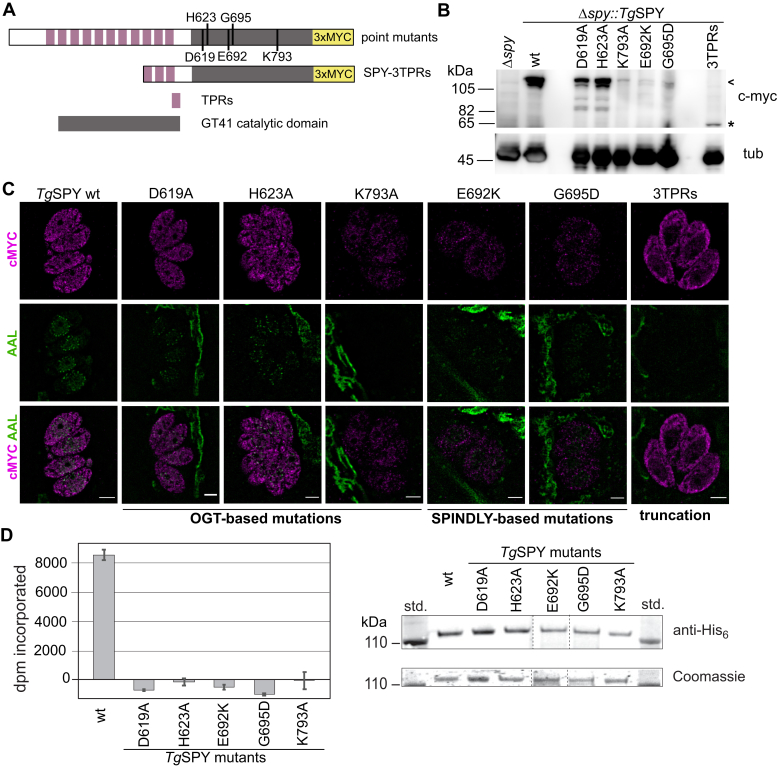
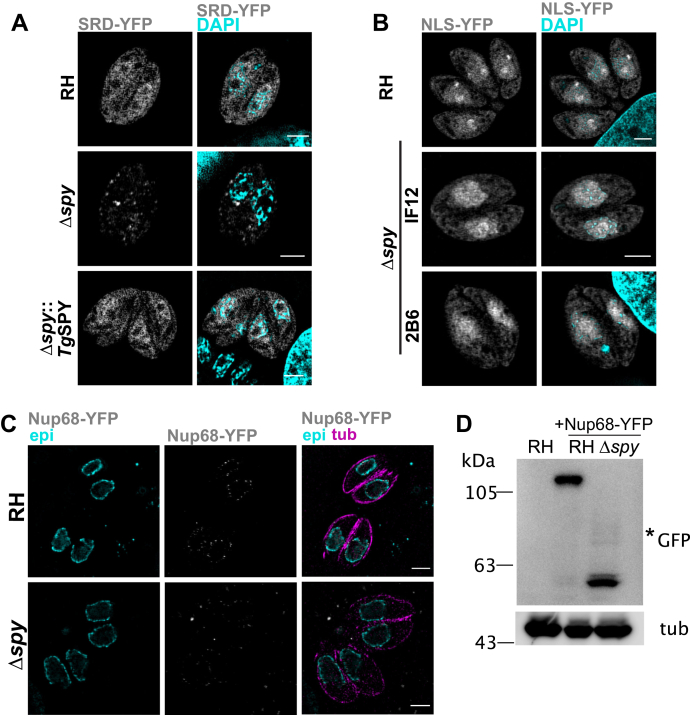
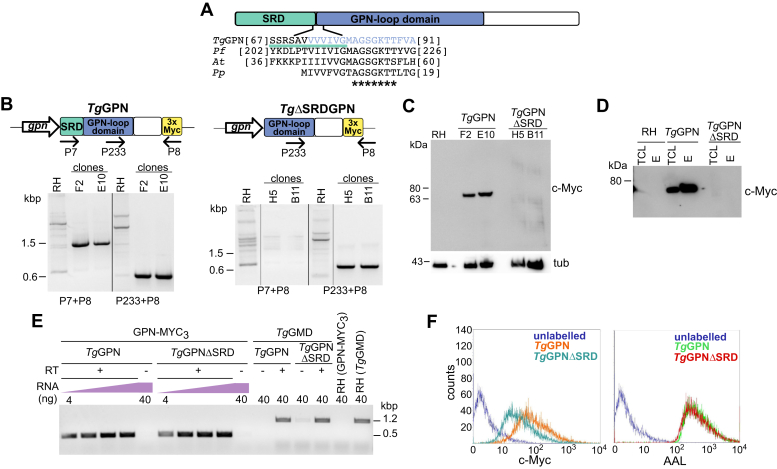
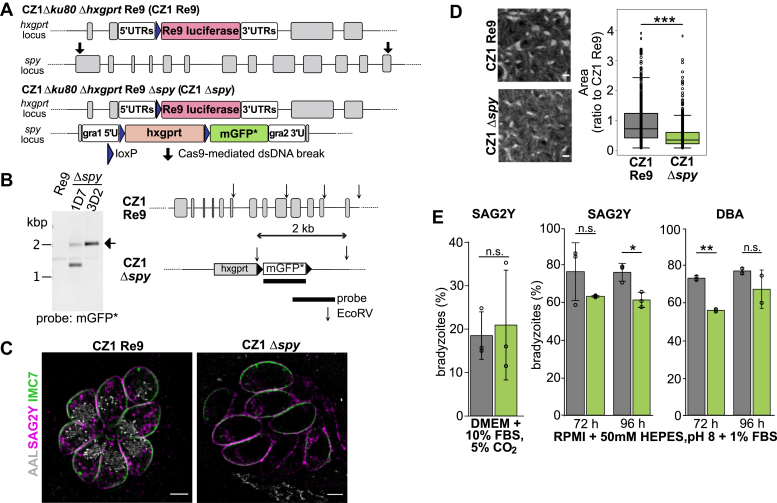
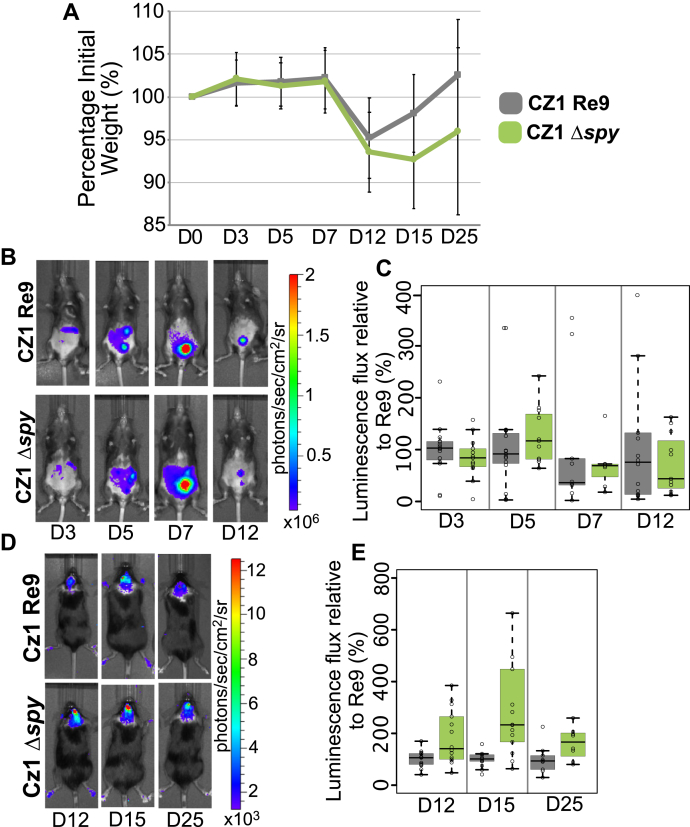
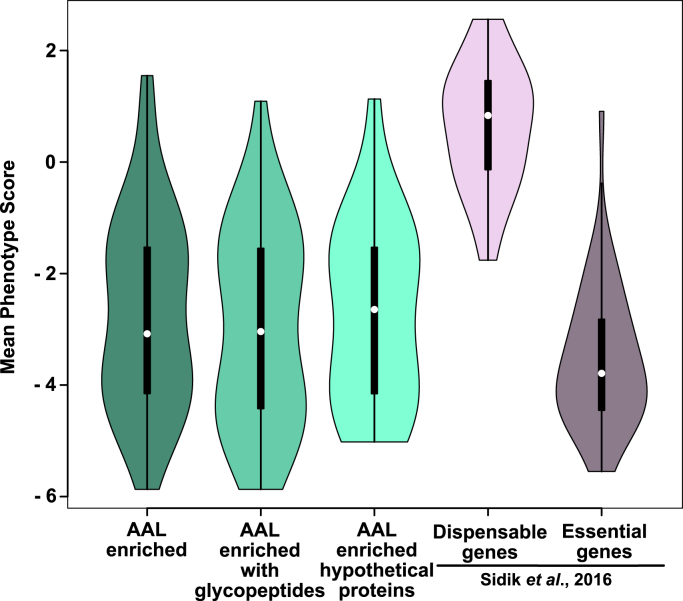
Once considered unusual, nucleocytoplasmic glycosylation is now recognized as a conserved feature of eukaryotes. While in animals, O-GlcNAc transferase (OGT) modifies thousands of intracellular proteins, the human pathogen Toxoplasma gondii transfers a different sugar, fucose, to proteins involved in transcription, mRNA processing, and signaling. Knockout experiments showed that TgSPY, an ortholog of plant SPINDLY and paralog of host OGT, is required for nuclear O-fucosylation. Here we verify that TgSPY is the nucleocytoplasmic O-fucosyltransferase (OFT) by 1) complementation with TgSPY-MYC3, 2) its functional dependence on amino acids critical for OGT activity, and 3) its ability to O-fucosylate itself and a model substrate and to specifically hydrolyze GDP-Fuc. While many of the endogenous proteins modified by O-Fuc are important for tachyzoite fitness, O-fucosylation by TgSPY is not essential. Growth of Δspy tachyzoites in fibroblasts is modestly affected, despite marked reductions in the levels of ectopically expressed proteins normally modified with O-fucose. Intact TgSPY-MYC3 localizes to the nucleus and cytoplasm, whereas catalytic mutants often displayed reduced abundance. Δspy tachyzoites of a luciferase-expressing type II strain exhibited infection kinetics in mice similar to wild-type but increased persistence in the chronic brain phase, potentially due to an imbalance of regulatory protein levels. The modest changes in parasite fitness in vitro and in mice, despite profound effects on reporter protein accumulation, and the characteristic punctate localization of O-fucosylated proteins suggest that TgSPY controls the levels of proteins to be held in reserve for response to novel stresses.
Keywords: Apicomplexa; Toxoplasma gondii; fucosyltransferase; glycosylation; nucleus; posttranslational modification; protein stability; structured illumination microscopy.
Copyright © 2020 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
-
- Torrey E.F., Yolken R.H. Toxoplasma oocysts as a public health problem. Trends Parasitol. 2013;29:380–384. - PubMed
-
- Lykins J., Li X., Levigne P., Zhou Y., El Bissati K., Clouser F., Wallon M., Morel F., Leahy K., El Mansouri B., Siddiqui M., Leong N., Michalowski M., Irwin E., Goodall P. Rapid, inexpensive, fingerstick, whole-blood, sensitive, specific, point-of-care test for anti-Toxoplasma antibodies. PLoS Negl. Trop. Dis. 2018;12:e0006536. - PMC - PubMed
-
- Guo M., Dubey J.P., Hill D., Buchanan R.L., Gamble H.R., Jones J.L., Pradhan A.K. Prevalence and risk factors for Toxoplasma gondii infection in meat animals and meat products destined for human consumption. J. Food Prot. 2015;78:457–476. - PubMed
-
- Pleyer U., Schlüter D., Mänz M. Ocular toxoplasmosis: recent aspects of pathophysiology and clinical implications. Ophthalmic Res. 2014;52:116–123. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
