

Emerging Functions of Actins and Actin Binding Proteins in Trypanosomatids
- PMID: 33163497
- PMCID: PMC7581878
- DOI: 10.3389/fcell.2020.587685
Emerging Functions of Actins and Actin Binding Proteins in Trypanosomatids
Abstract
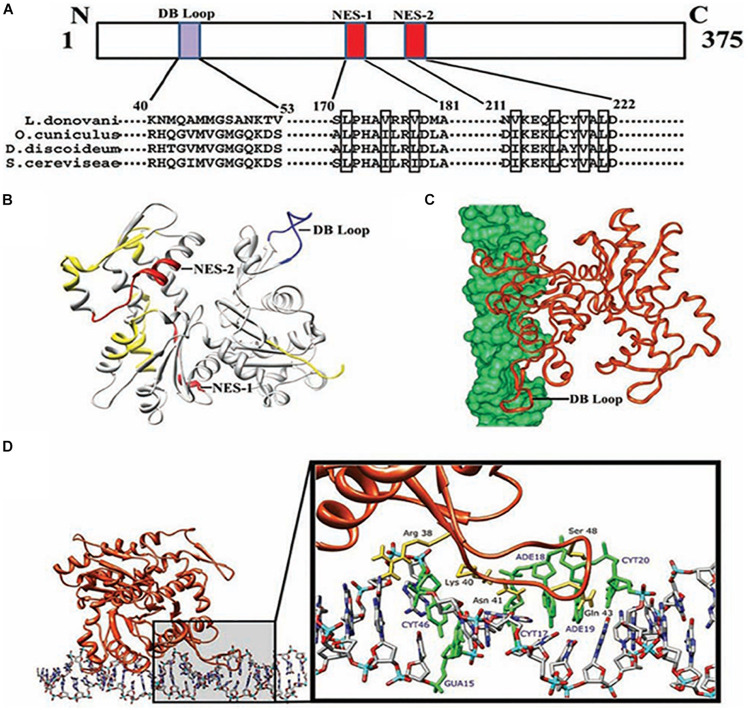
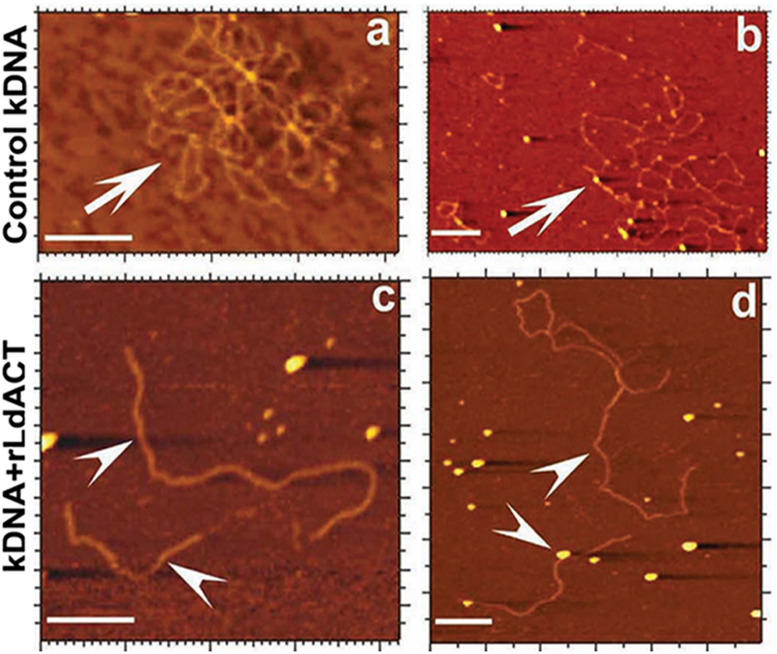
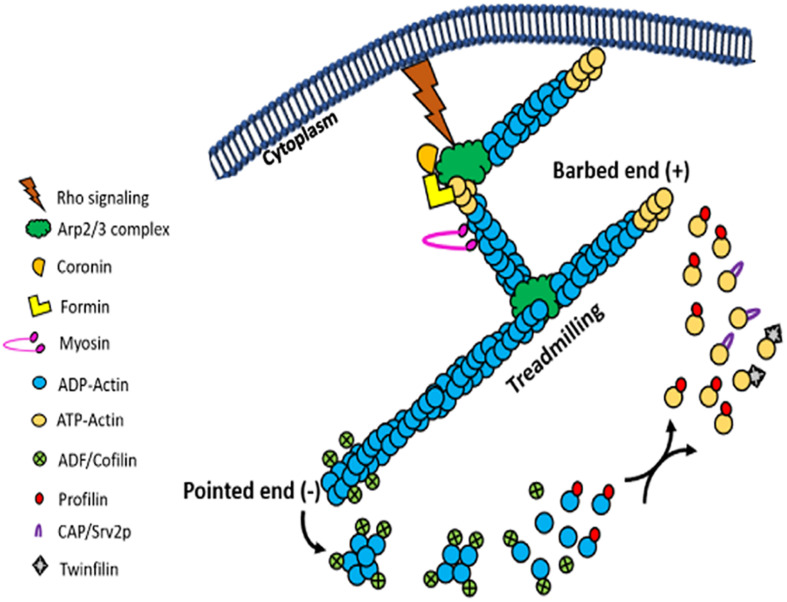
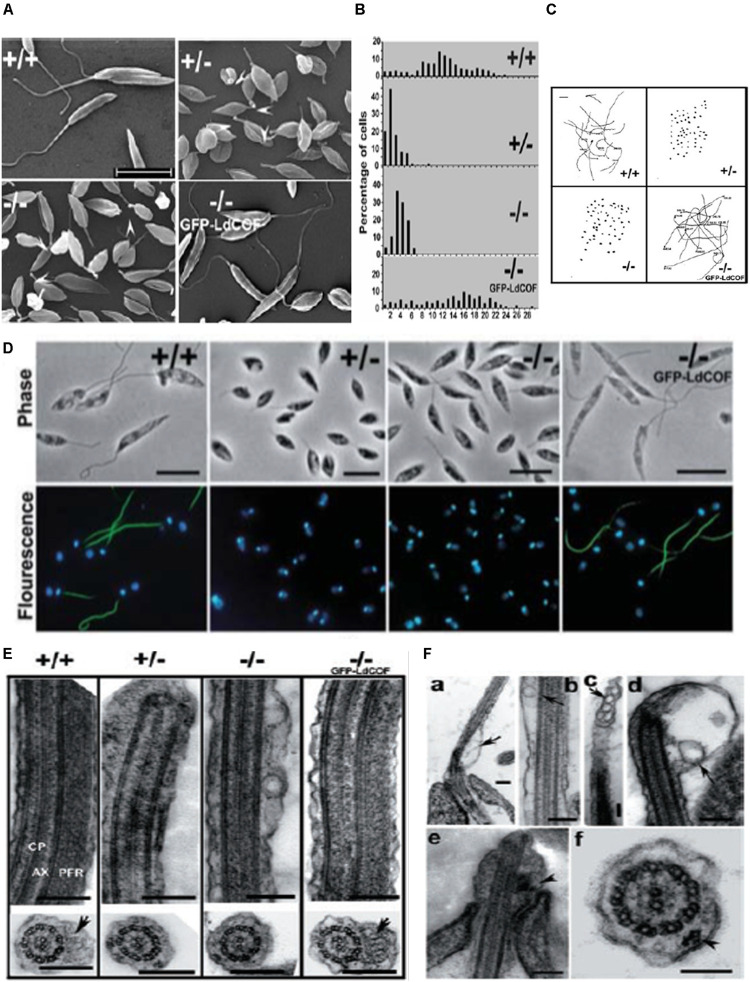
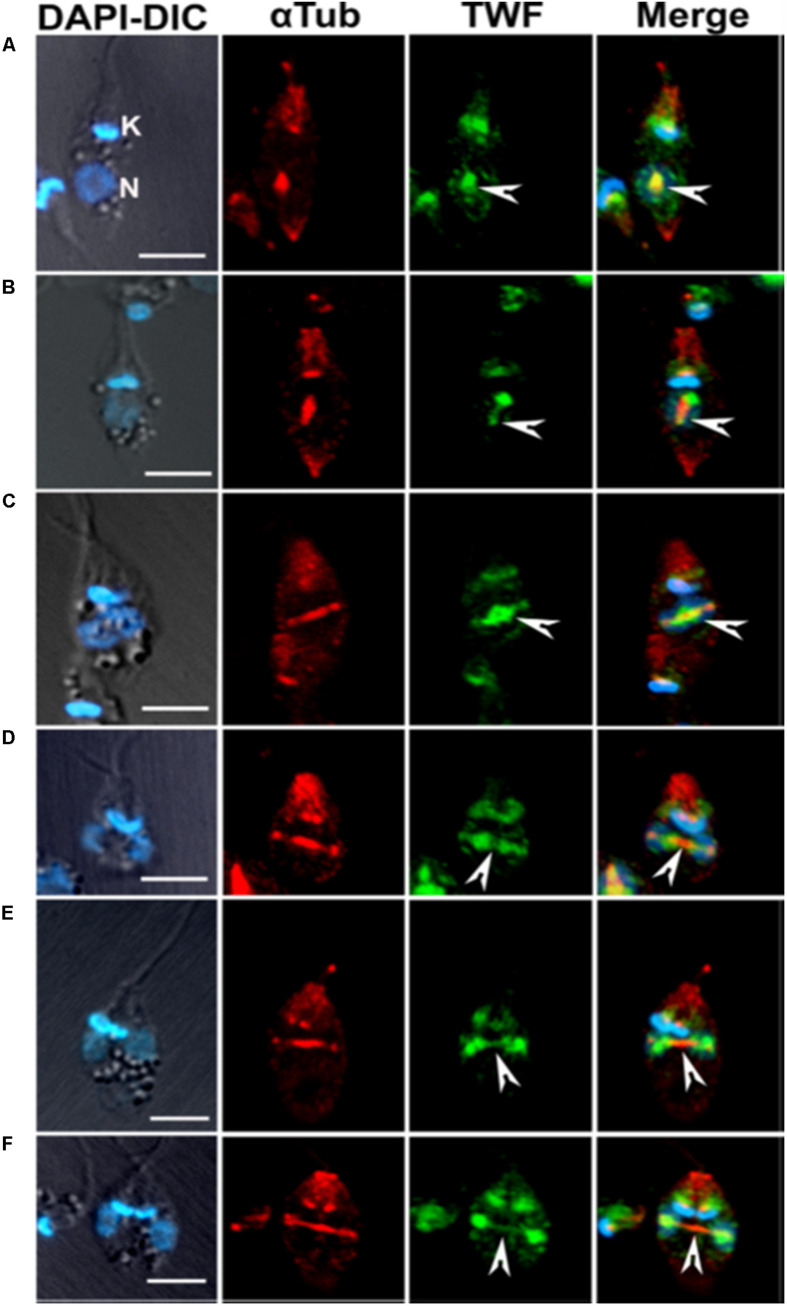
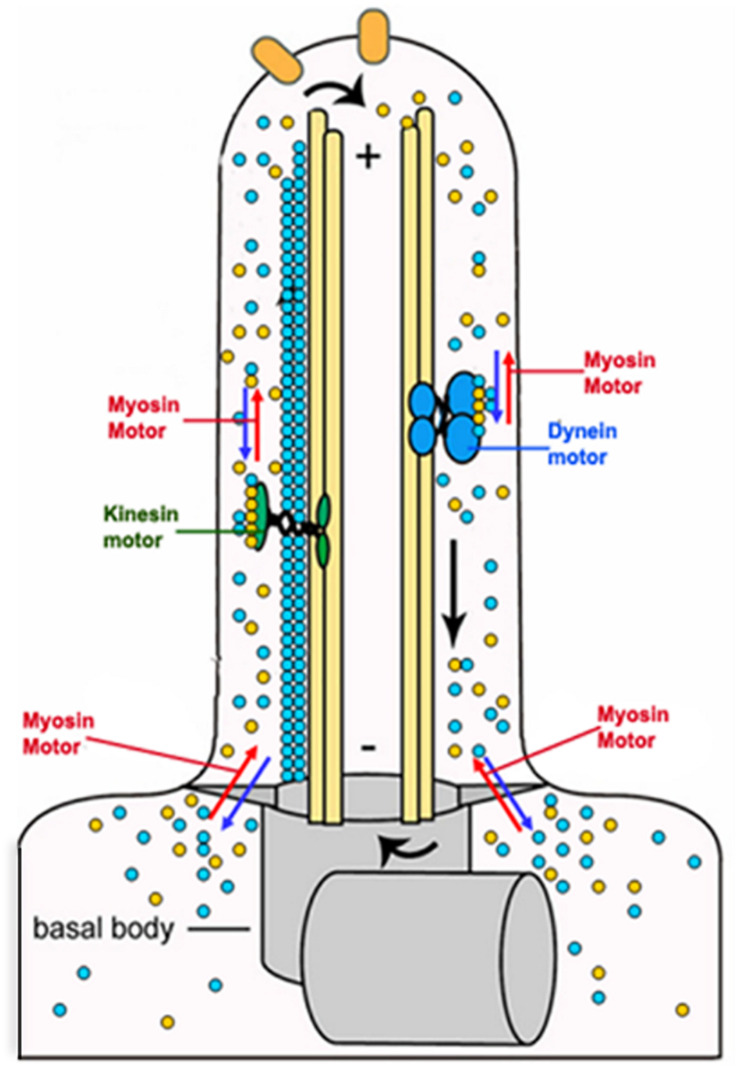
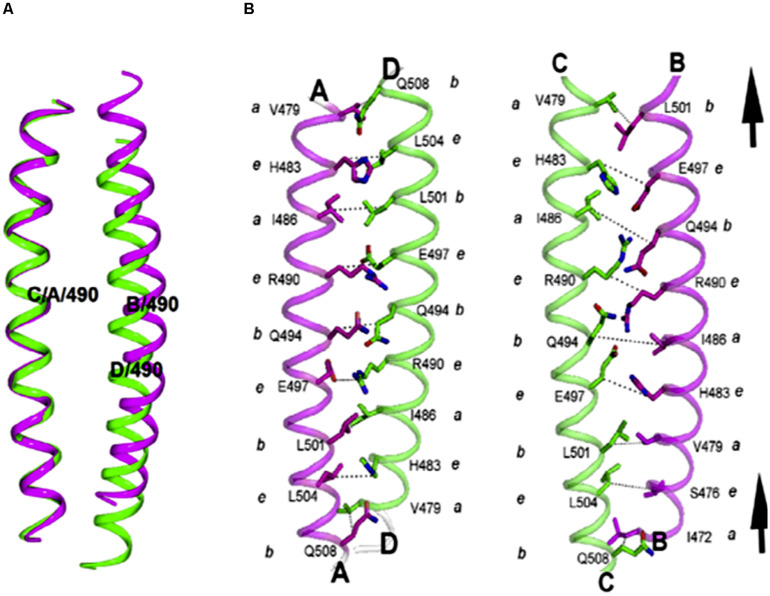
Actin is the major protein constituent of the cytoskeleton that performs wide range of cellular functions. It exists in monomeric and filamentous forms, dynamics of which is regulated by a large repertoire of actin binding proteins. However, not much was known about existence of these proteins in trypanosomatids, till the genome sequence data of three important organisms of this class, viz. Trypanosoma brucei, Trypanosoma cruzi and Leishmania major, became available. Here, we have reviewed most of the findings reported to date on the intracellular distribution, structure and functions of these proteins and based on them, we have hypothesized some of their functions. The major findings are as follows: (1) All the three organisms encode at least a set of ten actin binding proteins (profilin, twinfilin, ADF/cofilin, CAP/srv2, CAPz, coronin, two myosins, two formins) and one isoform of actin, except that T. cruzi encodes for three formins and several myosins along with four actins. (2) Actin 1 and a few actin binding proteins (ADF/cofilin, profilin, twinfilin, coronin and myosin13 in L. donovani; ADF/cofilin, profilin and myosin1 in T. brucei; profilin and myosin-F in T.cruzi) have been identified and characterized. (3) In all the three organisms, actin cytoskeleton has been shown to regulate endocytosis and intracellular trafficking. (4) Leishmania actin1 has been the most characterized protein among trypanosomatid actins. (5) This protein is localized to the cytoplasm as well as in the flagellum, nucleus and kinetoplast, and in vitro, it binds to DNA and displays scDNA relaxing and kDNA nicking activities. (6) The pure protein prefers to form bundles instead of thin filaments, and does not bind DNase1 or phalloidin. (7) Myosin13, myosin1 and myosin-F regulate endocytosis and intracellular trafficking, respectively, in Leishmania, T. brucei and T. cruzi. (8) Actin-dependent myosin13 motor is involved in dynamics and assembly of Leishmania flagellum. (9) Leishmania twinfilin localizes mostly to the nucleolus and coordinates karyokinesis by effecting splindle elongation and DNA synthesis. (10) Leishmania coronin binds and promotes actin filament formation and exists in tetrameric form rather than trimeric form, like other coronins. (11) Trypanosomatid profilins are essential for survival of all the three parasites.
Keywords: actin; actin binding proteins; functions; intracellular distribution; structure; tryanosomatids.
Copyright © 2020 Gupta, Ambaru and Bajaj.
Figures
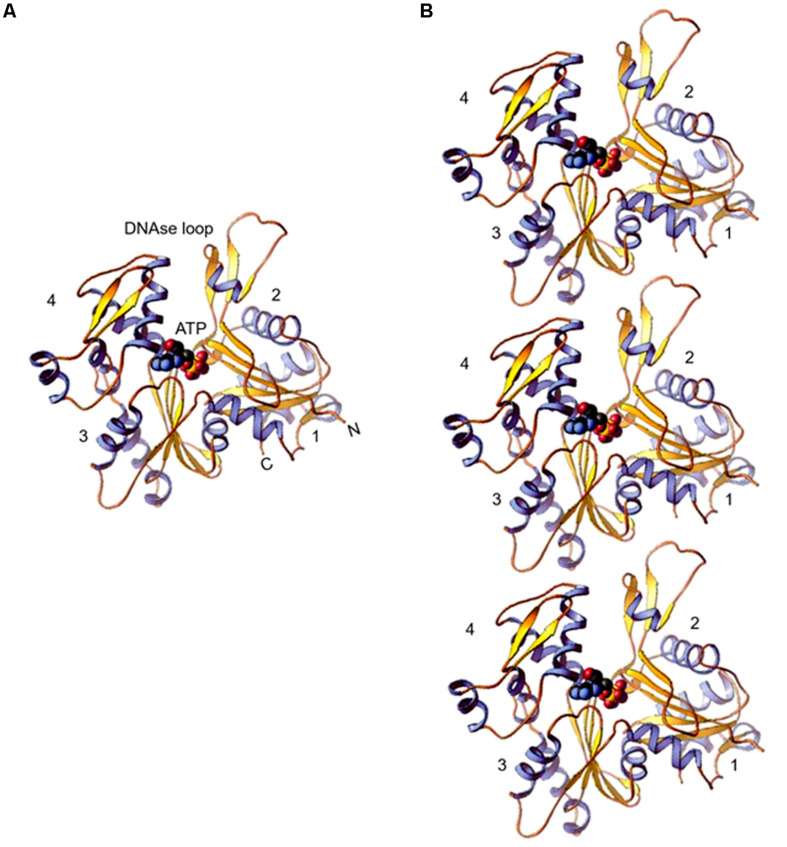
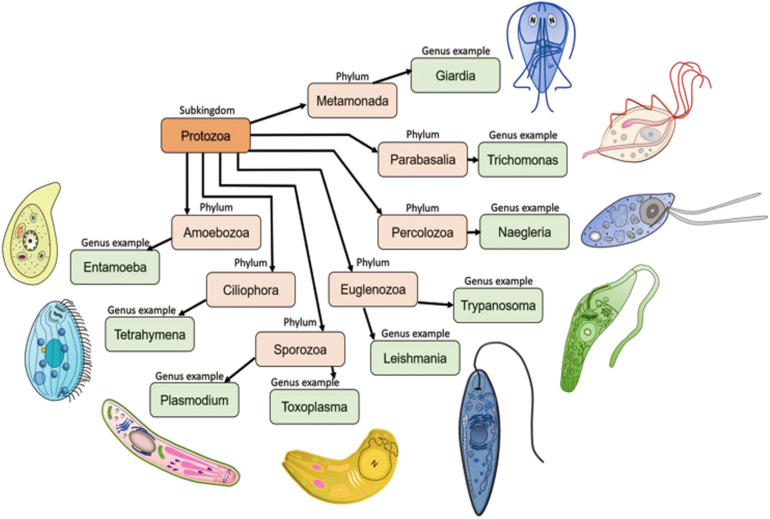
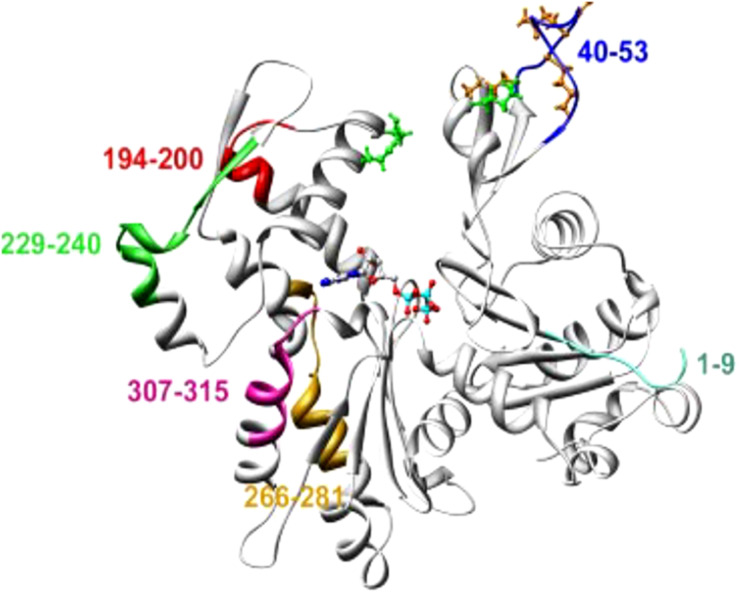
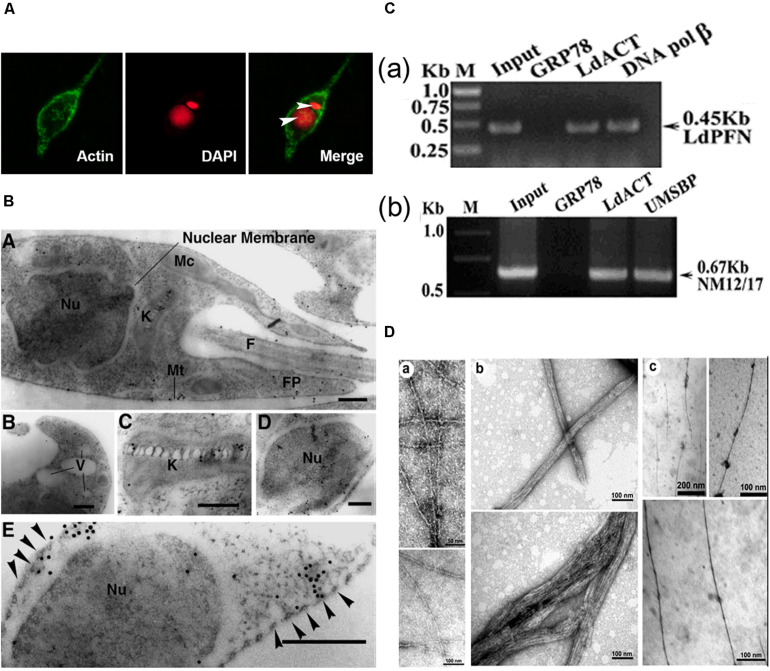
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous