

A frog with three sex chromosomes that co-mingle together in nature: Xenopus tropicalis has a degenerate W and a Y that evolved from a Z chromosome
- PMID: 33166278
- PMCID: PMC7652241
- DOI: 10.1371/journal.pgen.1009121
A frog with three sex chromosomes that co-mingle together in nature: Xenopus tropicalis has a degenerate W and a Y that evolved from a Z chromosome
Abstract
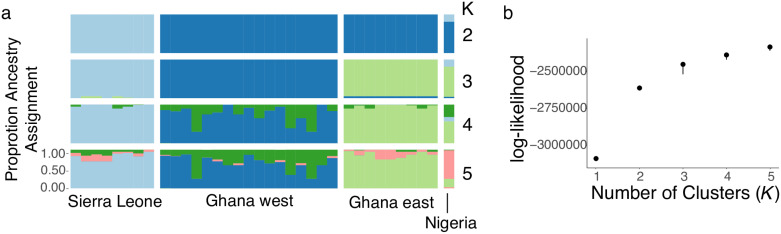
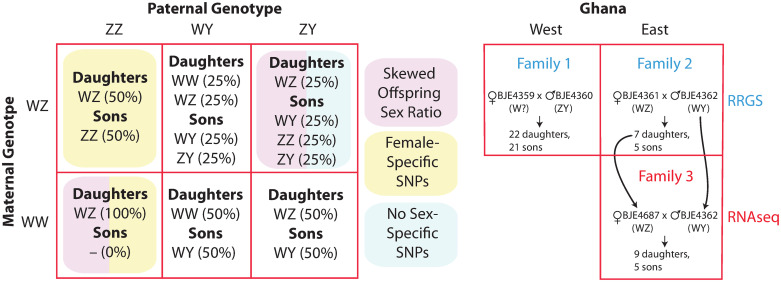
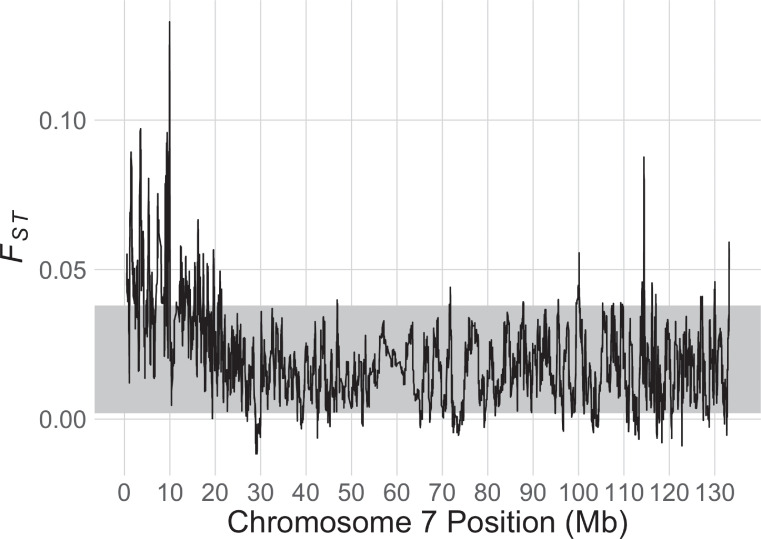
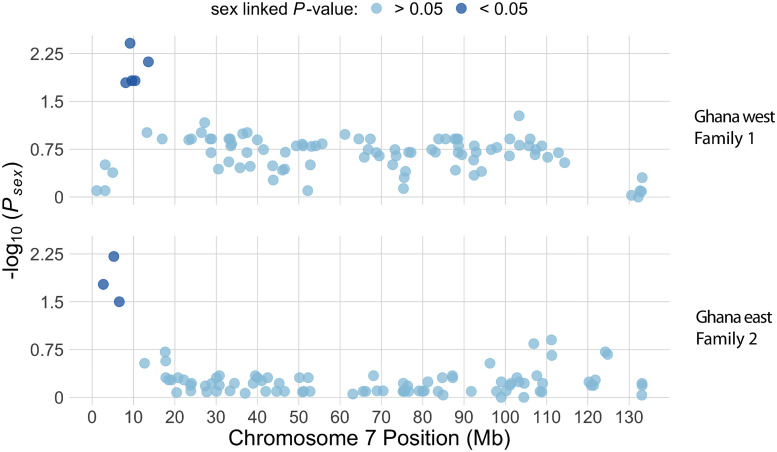
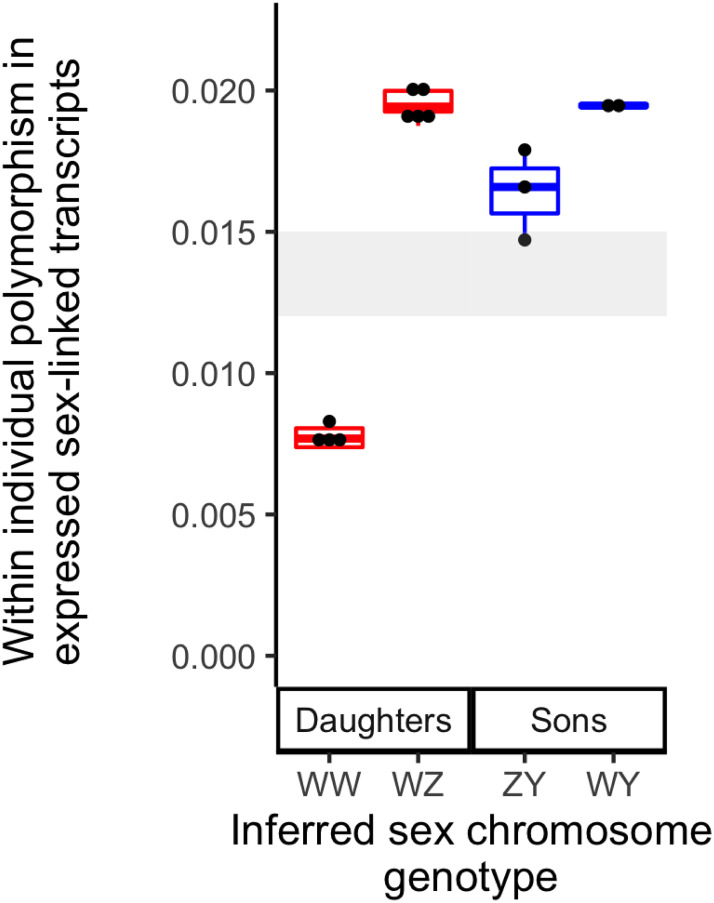
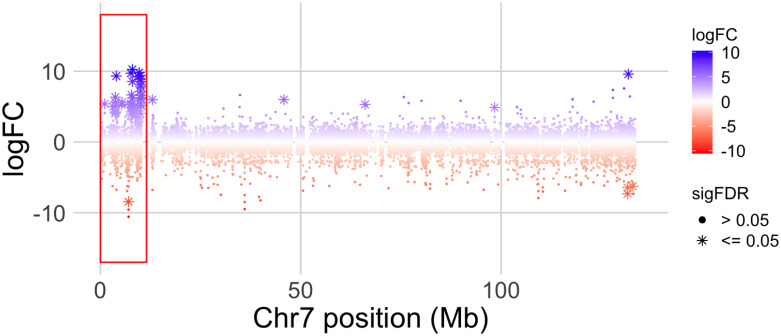
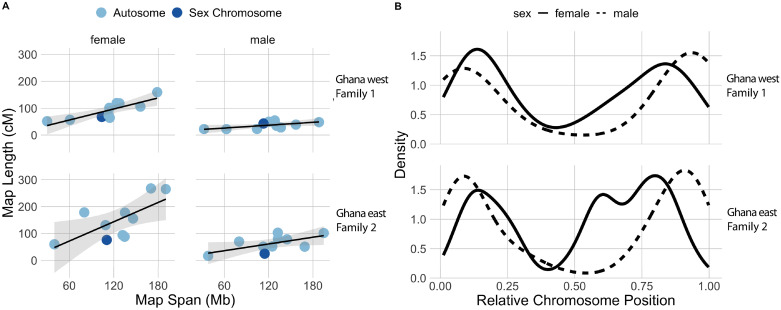
In many species, sexual differentiation is a vital prelude to reproduction, and disruption of this process can have severe fitness effects, including sterility. It is thus interesting that genetic systems governing sexual differentiation vary among-and even within-species. To understand these systems more, we investigated a rare example of a frog with three sex chromosomes: the Western clawed frog, Xenopus tropicalis. We demonstrate that natural populations from the western and eastern edges of Ghana have a young Y chromosome, and that a male-determining factor on this Y chromosome is in a very similar genomic location as a previously known female-determining factor on the W chromosome. Nucleotide polymorphism of expressed transcripts suggests genetic degeneration on the W chromosome, emergence of a new Y chromosome from an ancestral Z chromosome, and natural co-mingling of the W, Z, and Y chromosomes in the same population. Compared to the rest of the genome, a small sex-associated portion of the sex chromosomes has a 50-fold enrichment of transcripts with male-biased expression during early gonadal differentiation. Additionally, X. tropicalis has sex-differences in the rates and genomic locations of recombination events during gametogenesis that are similar to at least two other Xenopus species, which suggests that sex differences in recombination are genus-wide. These findings are consistent with theoretical expectations associated with recombination suppression on sex chromosomes, demonstrate that several characteristics of old and established sex chromosomes (e.g., nucleotide divergence, sex biased expression) can arise well before sex chromosomes become cytogenetically distinguished, and show how these characteristics can have lingering consequences that are carried forward through sex chromosome turnovers.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Matsubara K, Tarui H, Toriba M, Yamada K, Nishida-Umehara C, Agata K, et al. Evidence for different origin of sex chromosomes in snakes, birds, and mammals and step-wise differentiation of snake sex chromosomes. Proceedings of the National Academy of Sciences. 2006;103(48):18190–18195. 10.1073/pnas.0605274103 - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
