

Impact of subinhibitory concentrations of metronidazole on proteome of Clostridioides difficile strains with different levels of susceptibility
- PMID: 33166349
- PMCID: PMC7652271
- DOI: 10.1371/journal.pone.0241903
Impact of subinhibitory concentrations of metronidazole on proteome of Clostridioides difficile strains with different levels of susceptibility
Abstract
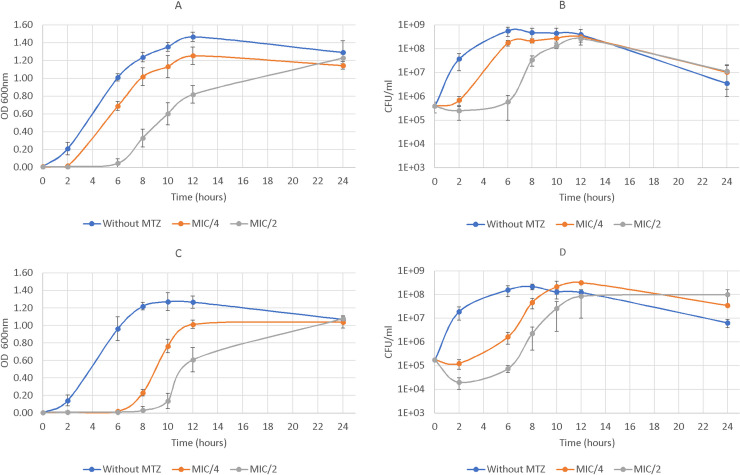
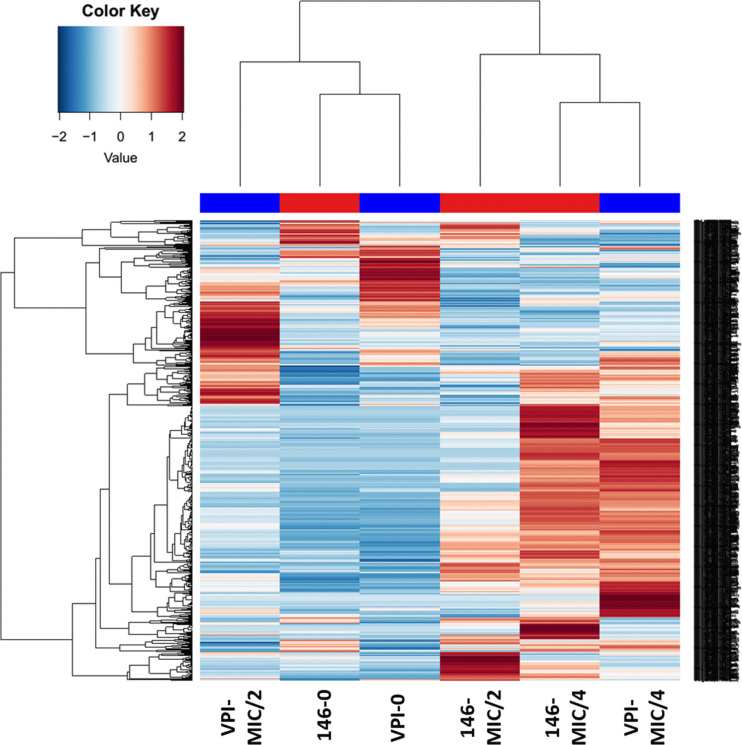
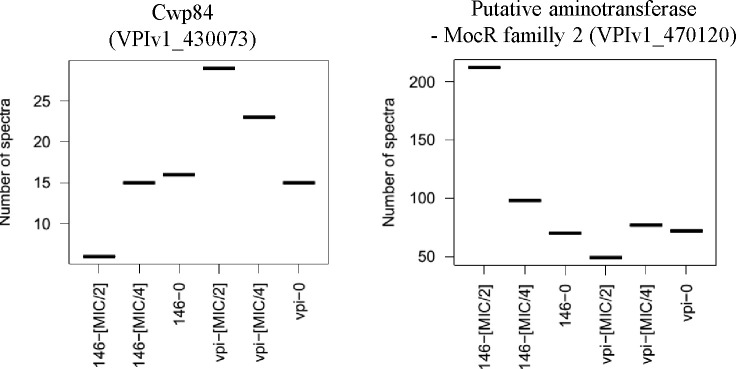
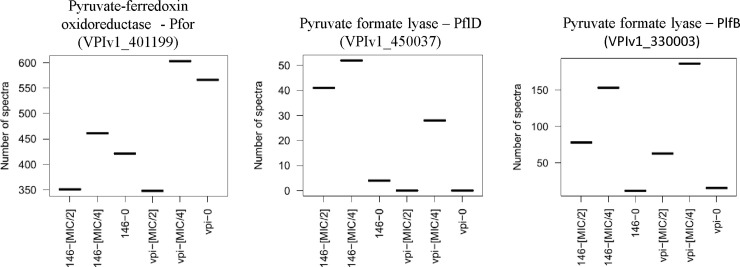
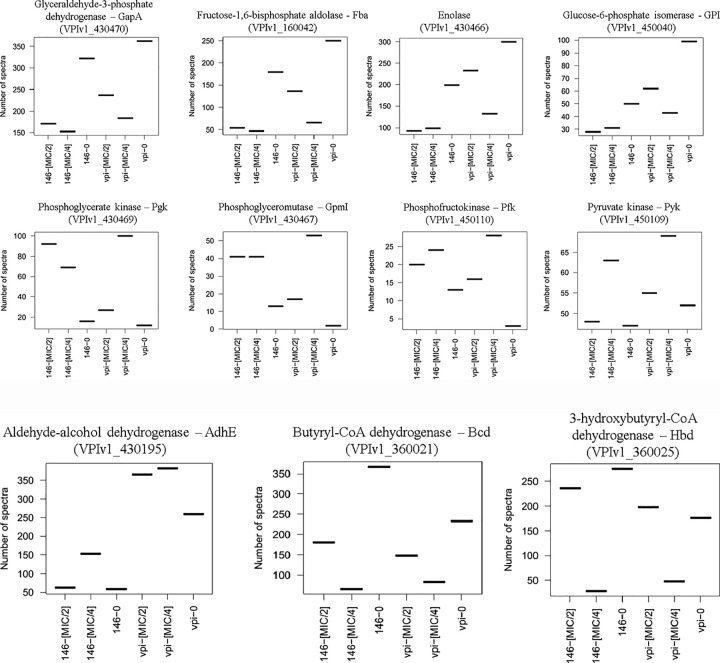
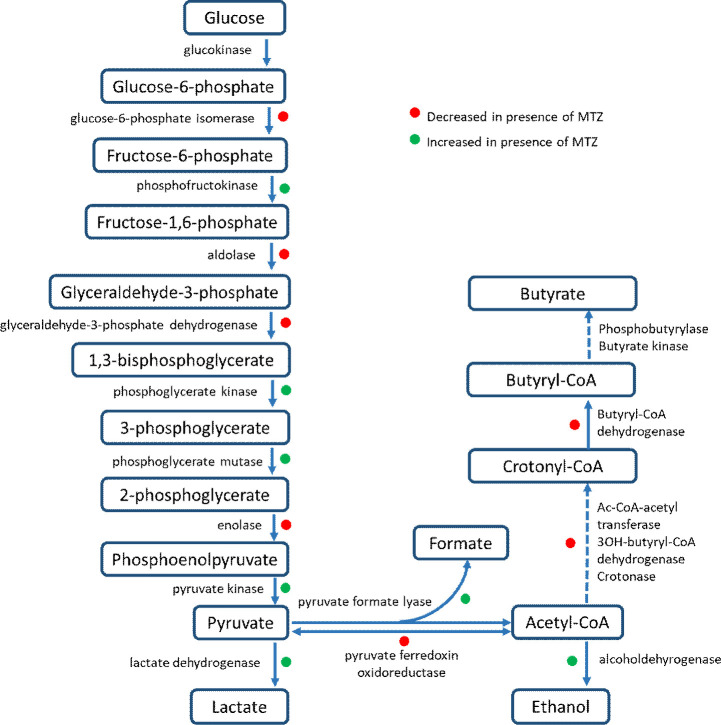
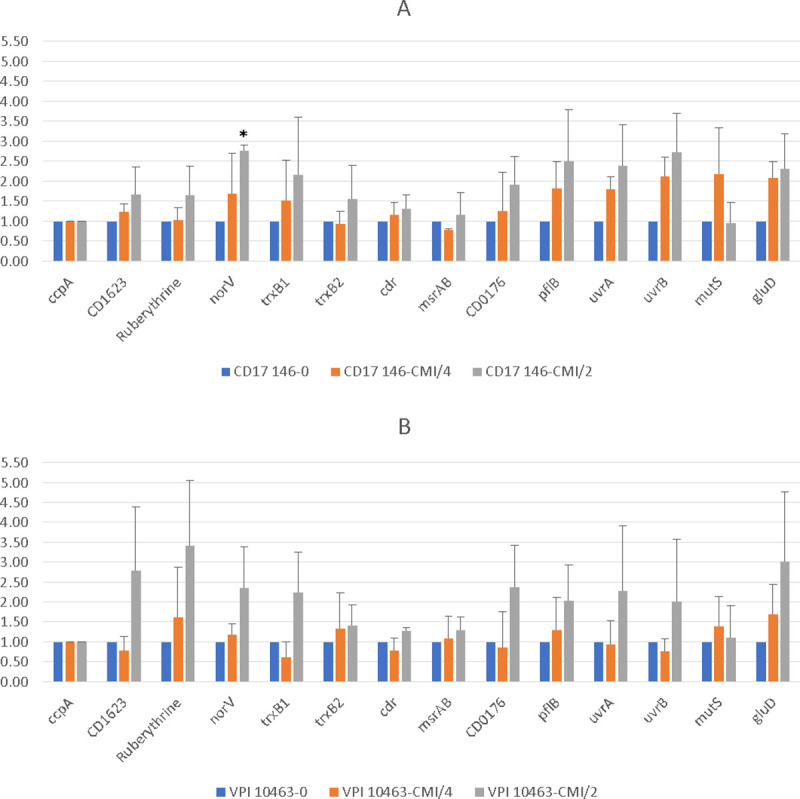
Clostridioides difficile is responsible for various intestinal symptoms from mild diarrhea to severe pseudomembranous colitis and is the primary cause of antibiotic-associated diarrhea in adults. Metronidazole was the first-line treatment for mild to moderate C. difficile infections for 30 years. However, clinical failure and recurrence rates of metronidazole is superior to oral vancomycin and metronidazole is now recommended only as an alternative to vancomycin or fidaxomicin, for an initial non-severe infection. The mechanisms of treatment failure and infection recurrence remain unclear. Given the poor fecal concentrations of metronidazole, the bacteria may be exposed to subinhibitory concentrations of metronidazole and develop adaptation strategy, which is likely to be the origin of an increase in treatment failures. In this study, a proteomic approach was used to analyze changes in the proteome of two strains with different levels of susceptibility to metronidazole in the presence of subinhibitory concentrations of this antibiotic. The two strains were grown to stationary phase: CD17-146, a clinical C. difficile isolate with reduced susceptibility to metronidazole, and VPI 10463, a metronidazole susceptible strain. Our study revealed that, whatever the strain, subinhibitory concentrations of metronidazole modified the amount of proteins involved in protein biosynthesis, glycolysis, and protection against stress induced by metronidazole, as well as in DNA repair. Several proteins involved in stress response are known to be synthesized under the control of Sigma factor B, which suggests a close link between Sigma factor B and metronidazole. Interestingly, impact of metronidazole on protein production for VPI 10463 strain differed from CD17-146 strain, for which the amount of two proteins involved in biofilm formation of CD17-146 were modified by metronidazole.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- McDonald LC, Gerding DN, Johnson S, Bakken JS, Carroll KC, Coffin SE, et al. Clinical Practice Guidelines for Clostridium difficile Infection in Adults and Children: 2017 Update by the Infectious Diseases Society of America (IDSA) and Society for Healthcare Epidemiology of America (SHEA). Clin Infect Dis Off Publ Infect Dis Soc Am 2018;66:e1–48. 10.1093/cid/cix1085. - DOI - PMC - PubMed
-
- Johnson S, Louie TJ, Gerding DN, Cornely OA, Chasan-Taber S, Fitts D, et al. Vancomycin, metronidazole, or tolevamer for Clostridium difficile infection: results from two multinational, randomized, controlled trials. Clin Infect Dis Off Publ Infect Dis Soc Am 2014;59:345–54. 10.1093/cid/ciu313 - DOI - PubMed
-
- Cornely OA, Crook DW, Esposito R, Poirier A, Somero MS, Weiss K, et al. Fidaxomicin versus vancomycin for infection with Clostridium difficile in Europe, Canada, and the USA: a double-blind, non-inferiority, randomised controlled trial. Lancet Infect Dis 2012;12:281–9. 10.1016/S1473-3099(11)70374-7 - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
