

Identification of a brainstem locus that inhibits tumor necrosis factor
- PMID: 33168718
- PMCID: PMC7703602
- DOI: 10.1073/pnas.2008213117
Identification of a brainstem locus that inhibits tumor necrosis factor
Abstract
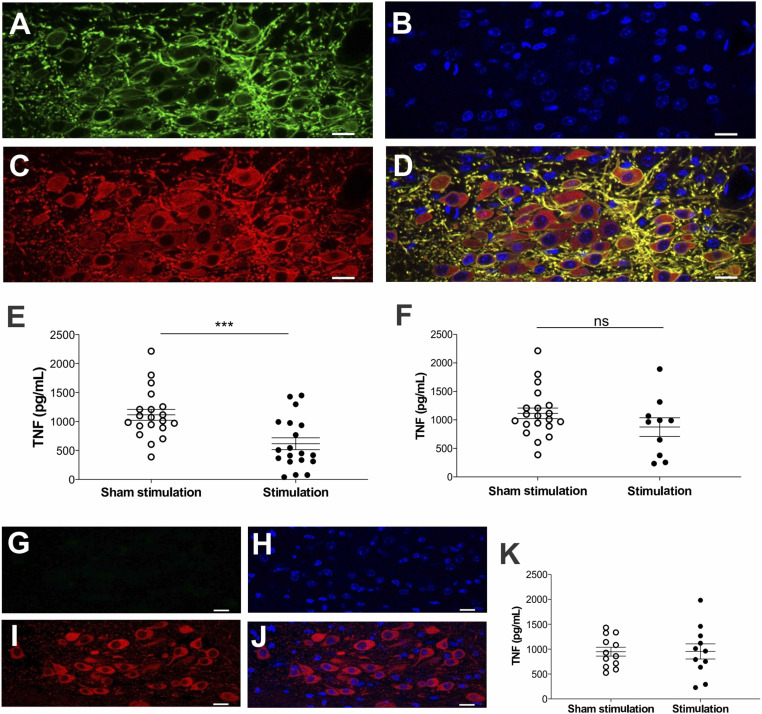
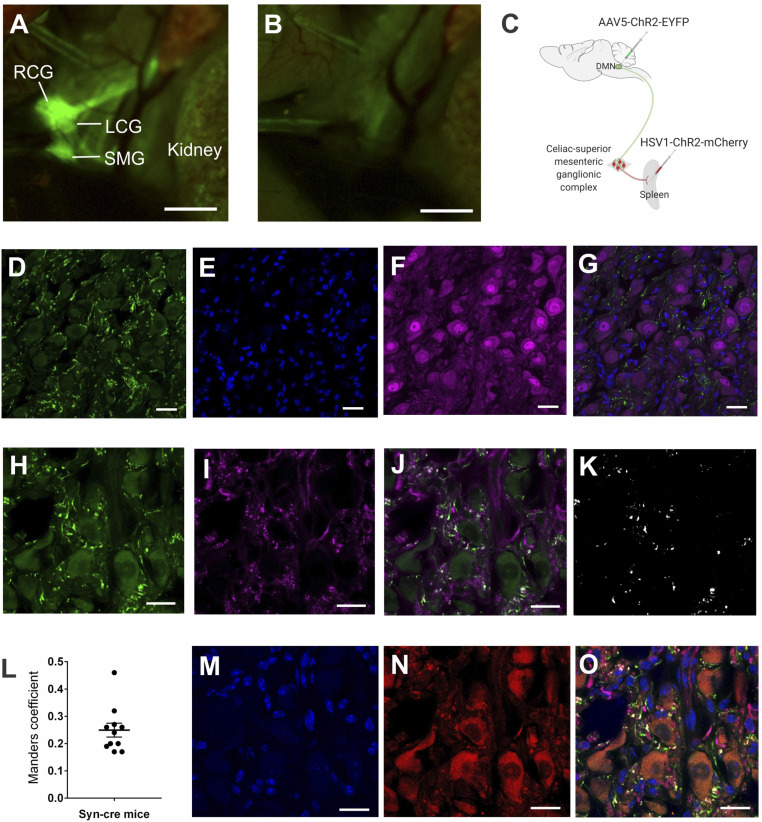
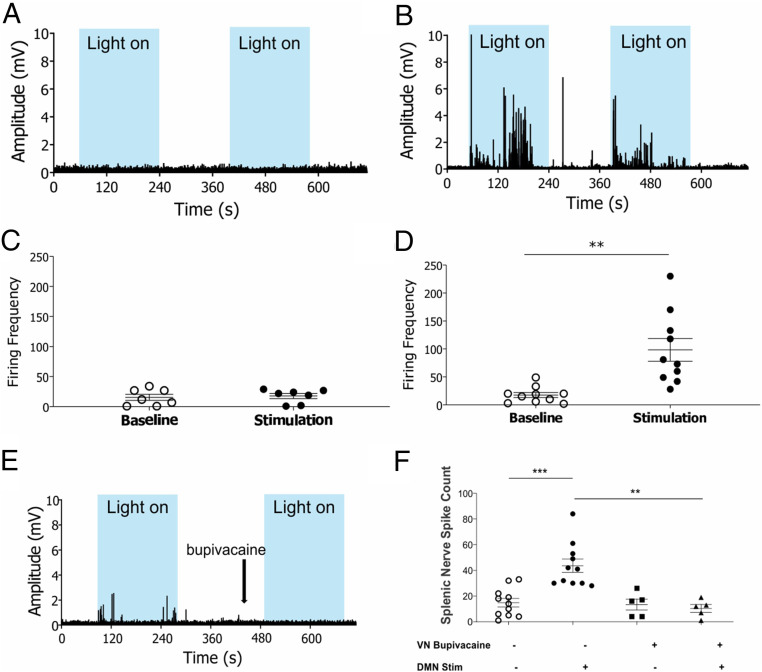
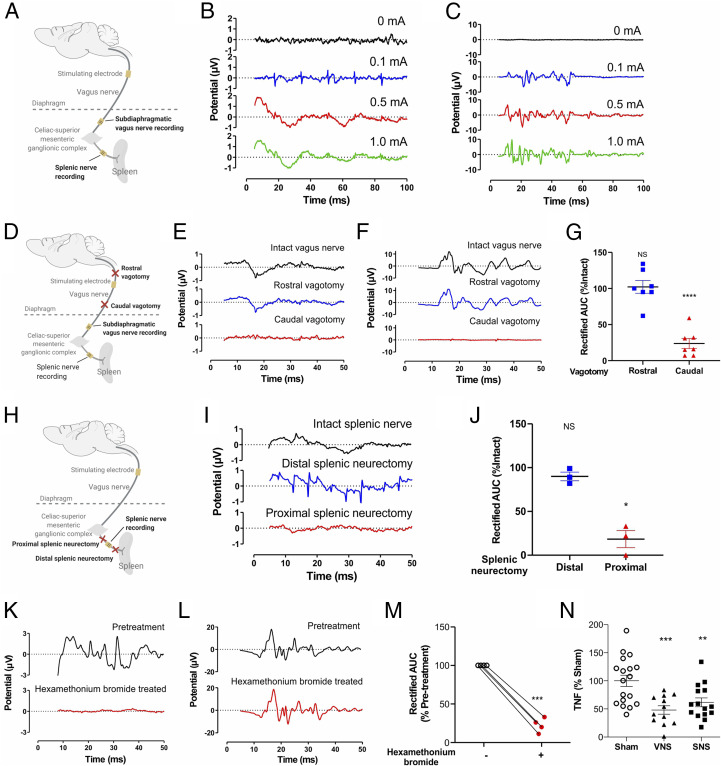
In the brain, compact clusters of neuron cell bodies, termed nuclei, are essential for maintaining parameters of host physiology within a narrow range optimal for health. Neurons residing in the brainstem dorsal motor nucleus (DMN) project in the vagus nerve to communicate with the lungs, liver, gastrointestinal tract, and other organs. Vagus nerve-mediated reflexes also control immune system responses to infection and injury by inhibiting the production of tumor necrosis factor (TNF) and other cytokines in the spleen, although the function of DMN neurons in regulating TNF release is not known. Here, optogenetics and functional mapping reveal cholinergic neurons in the DMN, which project to the celiac-superior mesenteric ganglia, significantly increase splenic nerve activity and inhibit TNF production. Efferent vagus nerve fibers terminating in the celiac-superior mesenteric ganglia form varicose-like structures surrounding individual nerve cell bodies innervating the spleen. Selective optogenetic activation of DMN cholinergic neurons or electrical activation of the cervical vagus nerve evokes action potentials in the splenic nerve. Pharmacological blockade and surgical transection of the vagus nerve inhibit vagus nerve-evoked splenic nerve responses. These results indicate that cholinergic neurons residing in the brainstem DMN control TNF production, revealing a role for brainstem coordination of immunity.
Keywords: TNF; cytokines; dorsal motor nucleus; inflammatory reflex; vagus nerve.
Copyright © 2020 the Author(s). Published by PNAS.
Conflict of interest statement
Competing interest statement: A.M.K., V.A.P., S.S.C. and K.J.T. have filed a patent application relevant to this work and have assigned their rights to the Feinstein Institutes for Medical Research. Y.A.L. is an employee of SetPoint Medical.
Figures
References
-
- Laughton W. B., Powley T. L., Localization of efferent function in the dorsal motor nucleus of the vagus. Am. J. Physiol. 252, R13–R25 (1987). - PubMed
-
- Ionescu E., Rohner-Jeanrenaud F., Berthoud H. R., Jeanrenaud B., Increases in plasma insulin levels in response to electrical stimulation of the dorsal motor nucleus of the vagus nerve. Endocrinology 112, 904–910 (1983). - PubMed
-
- Geis G. S., Wurster R. D., Cardiac responses during stimulation of the dorsal motor nucleus and nucleus ambiguus in the cat. Circ. Res. 46, 606–611 (1980). - PubMed
-
- Montgomery J. P., Patterson P. H., Behavioral stress and tumor progression. Anticancer Res. 26, 1189–1192 (2006). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
