

TBK1 regulates regeneration of pancreatic β-cells
- PMID: 33168920
- PMCID: PMC7653919
- DOI: 10.1038/s41598-020-76600-6
TBK1 regulates regeneration of pancreatic β-cells
Abstract
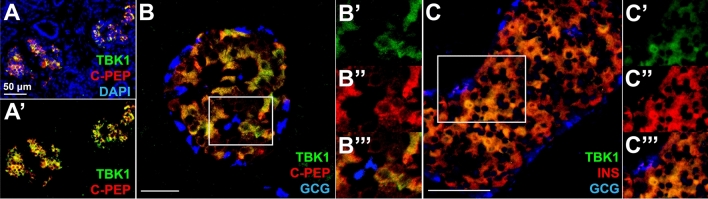
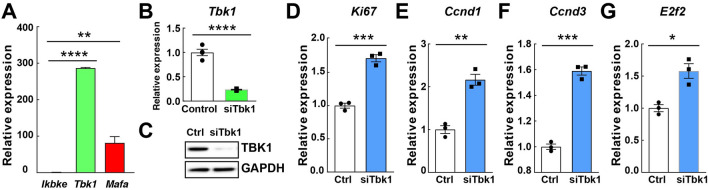
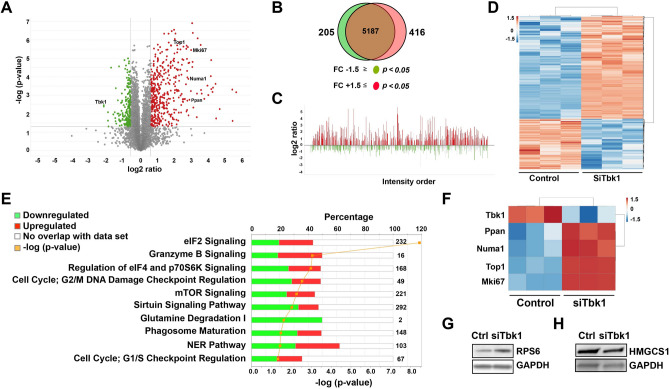
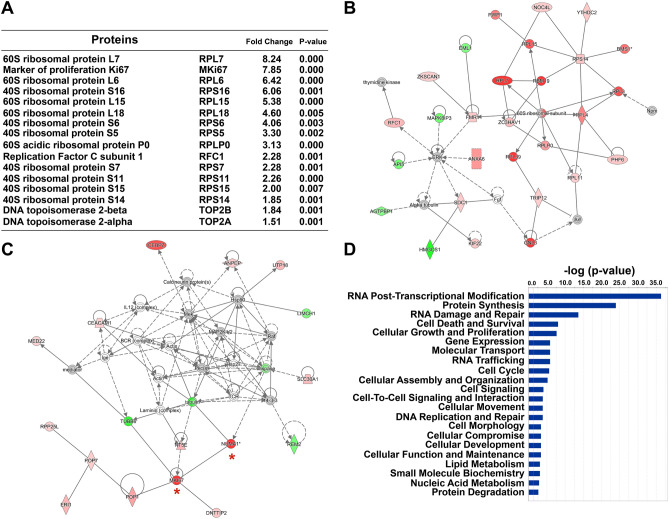
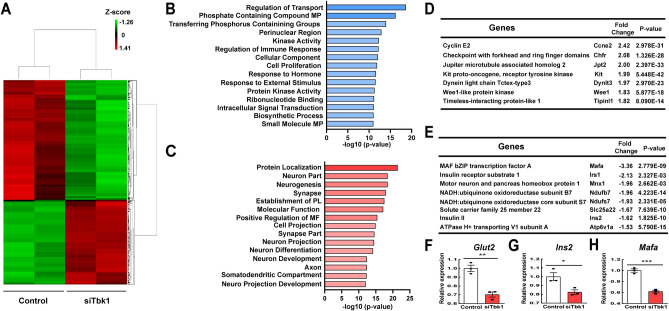
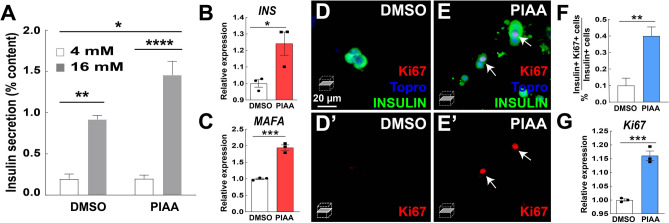
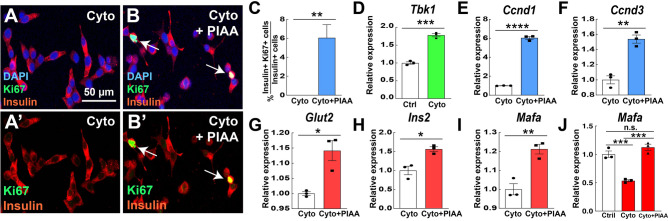
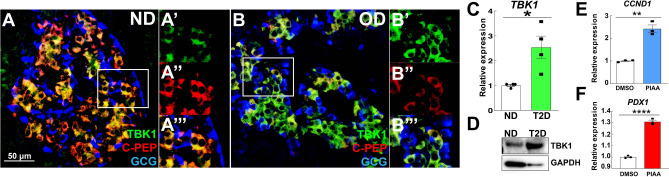
Small-molecule inhibitors of non-canonical IκB kinases TANK-binding kinase 1 (TBK1) and IκB kinase ε (IKKε) have shown to stimulate β-cell regeneration in multiple species. Here we demonstrate that TBK1 is predominantly expressed in β-cells in mammalian islets. Proteomic and transcriptome analyses revealed that genetic silencing of TBK1 increased expression of proteins and genes essential for cell proliferation in INS-1 832/13 rat β-cells. Conversely, TBK1 overexpression decreased sensitivity of β-cells to the elevation of cyclic AMP (cAMP) levels and reduced proliferation of β-cells in a manner dependent on the activity of cAMP-hydrolyzing phosphodiesterase 3 (PDE3). While the mitogenic effect of (E)3-(3-phenylbenzo[c]isoxazol-5-yl)acrylic acid (PIAA) is derived from inhibition of TBK1, PIAA augmented glucose-stimulated insulin secretion (GSIS) and expression of β-cell differentiation and proliferation markers in human embryonic stem cell (hESC)-derived β-cells and human islets. TBK1 expression was increased in β-cells upon diabetogenic insults, including in human type 2 diabetic islets. PIAA enhanced expression of cell cycle control molecules and β-cell differentiation markers upon diabetogenic challenges, and accelerated restoration of functional β-cells in streptozotocin (STZ)-induced diabetic mice. Altogether, these data suggest the critical function of TBK1 as a β-cell autonomous replication barrier and present PIAA as a valid therapeutic strategy augmenting functional β-cells.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
