

Extracellular Vesicles Released by Enterovirus-Infected EndoC-βH1 Cells Mediate Non-Lytic Viral Spread
- PMID: 33171580
- PMCID: PMC7695210
- DOI: 10.3390/microorganisms8111753
Extracellular Vesicles Released by Enterovirus-Infected EndoC-βH1 Cells Mediate Non-Lytic Viral Spread
Abstract
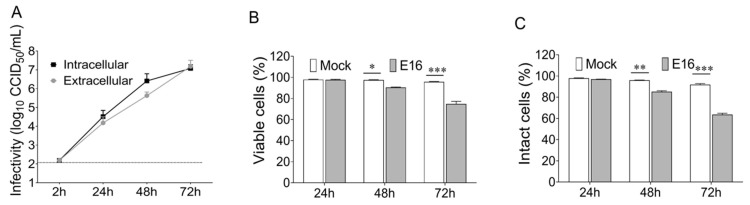
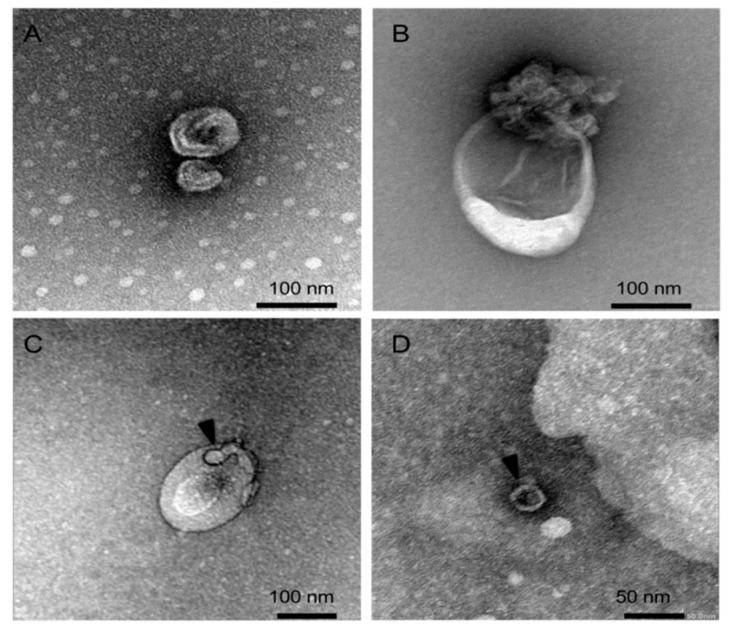
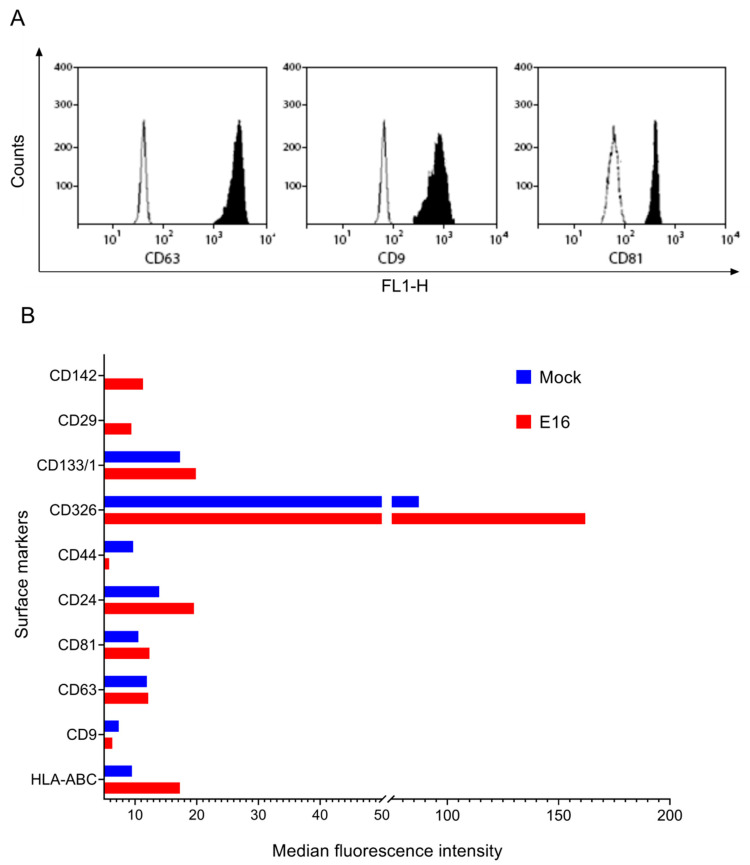
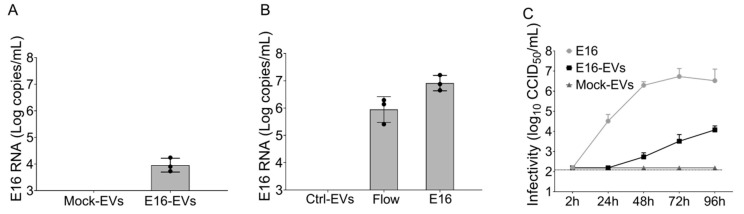
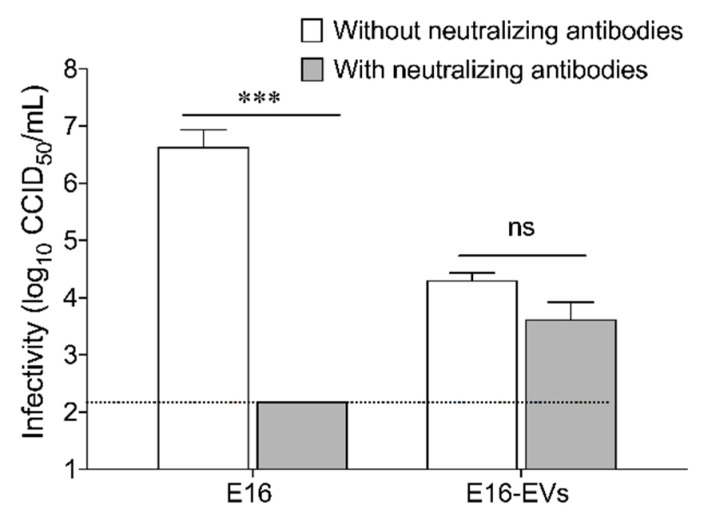
While human enteroviruses are generally regarded as a lytic virus, and persistent non-cytolytic enterovirus infection in pancreatic beta cells has been suspected of playing a role in type 1 diabetes pathogenesis. However, it is still unclear how enteroviruses could exit the pancreatic beta cell in a non-lytic manner. This study aimed to investigate the role of beta cell-derived extracellular vesicles (EVs) in the non-lytic enteroviral spread and infection. Size-exclusion chromatography and antibody-based immunoaffinity purification were used to isolate EVs from echovirus 16-infected human beta EndoC-βH1 cells. EVs were then characterized using transmission electron microscopy and Multiplex Bead-Based Flow Cytometry Assay. Virus production and release were quantified by 50% cell culture infectious dose (CCID50) assay and qRT-PCR. Our results showed that EVs from echovirus 16-infected EndoC-βH1 cells harbor infectious viruses and promote their spread during the pre-lytic phase of infection. Furthermore, the EVs-mediated infection was not inhibited by virus-specific neutralizing antibodies. In summary, this study demonstrated that enteroviruses could exit beta cells non-lytically within infectious EVs, thereby thwarting the access of neutralizing antibodies to viral particles. These data suggest that enterovirus transmission through EVs may contribute to viral dissemination and immune evasion in persistently infected beta cells.
Keywords: beta cells; enterovirus; exosomes; extracellular vesicles; type 1 diabetes; virus spread.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
References
-
- Pallansch M., Roos R. Enteroviruses: Polioviruses, coxsackieviruses, echoviruses and newer enteroviruses. In: Knipe D.M., Howley P.M., editors. Fields Virology. 5th ed. Lippincott Williams & Wilkins; Philadelphia, PA, USA: 2006. pp. 895–910.
-
- Hsu N.Y., Ilnytska O., Belov G., Santiana M., Chen Y.H., Takvorian P.M., Pau C., van der Schaar H., Kaushik-Basu N., Balla T., et al. Viral reorganization of the secretory pathway generates distinct organelles for RNA replication. Cell. 2010;141:799–811. doi: 10.1016/j.cell.2010.03.050. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
