

Nanotherapeutic Modulation of Human Neural Cells and Glioblastoma in Organoids and Monocultures
- PMID: 33171886
- PMCID: PMC7695149
- DOI: 10.3390/cells9112434
Nanotherapeutic Modulation of Human Neural Cells and Glioblastoma in Organoids and Monocultures
Abstract
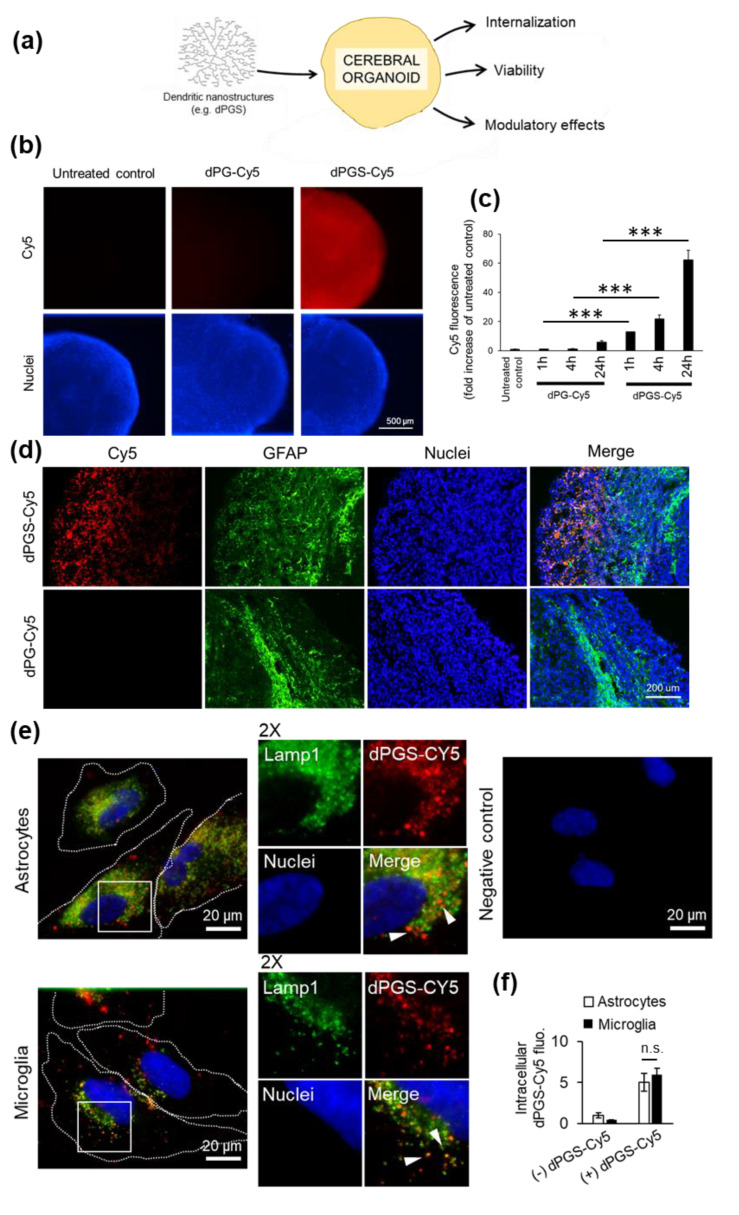
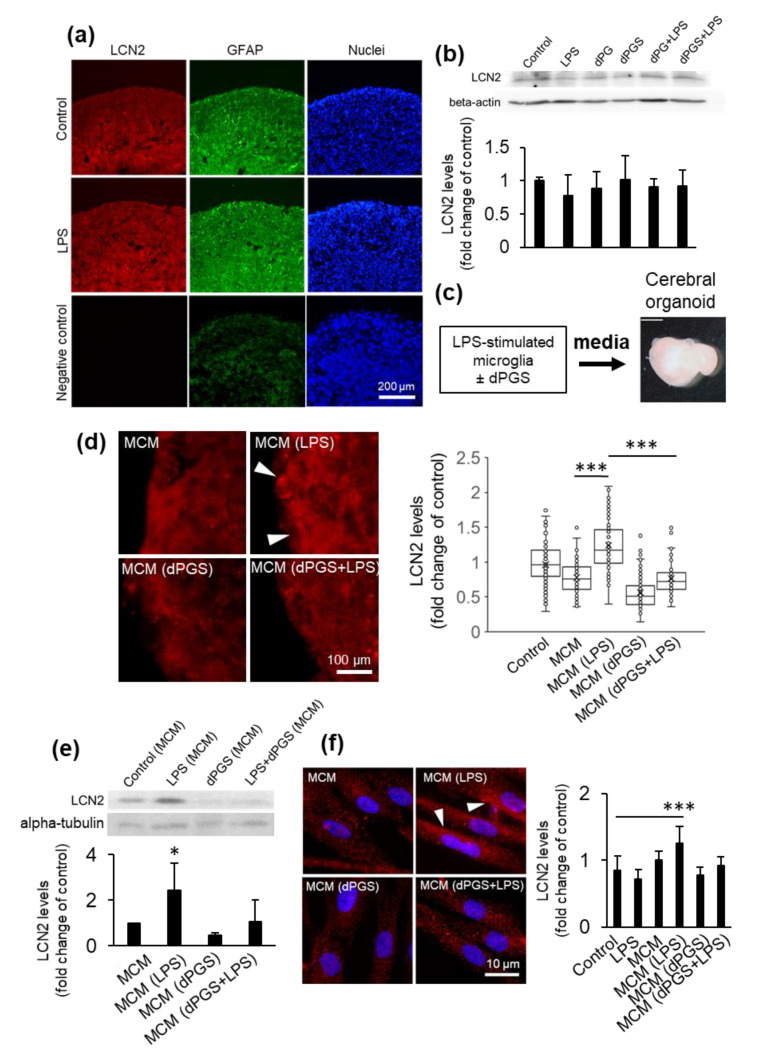
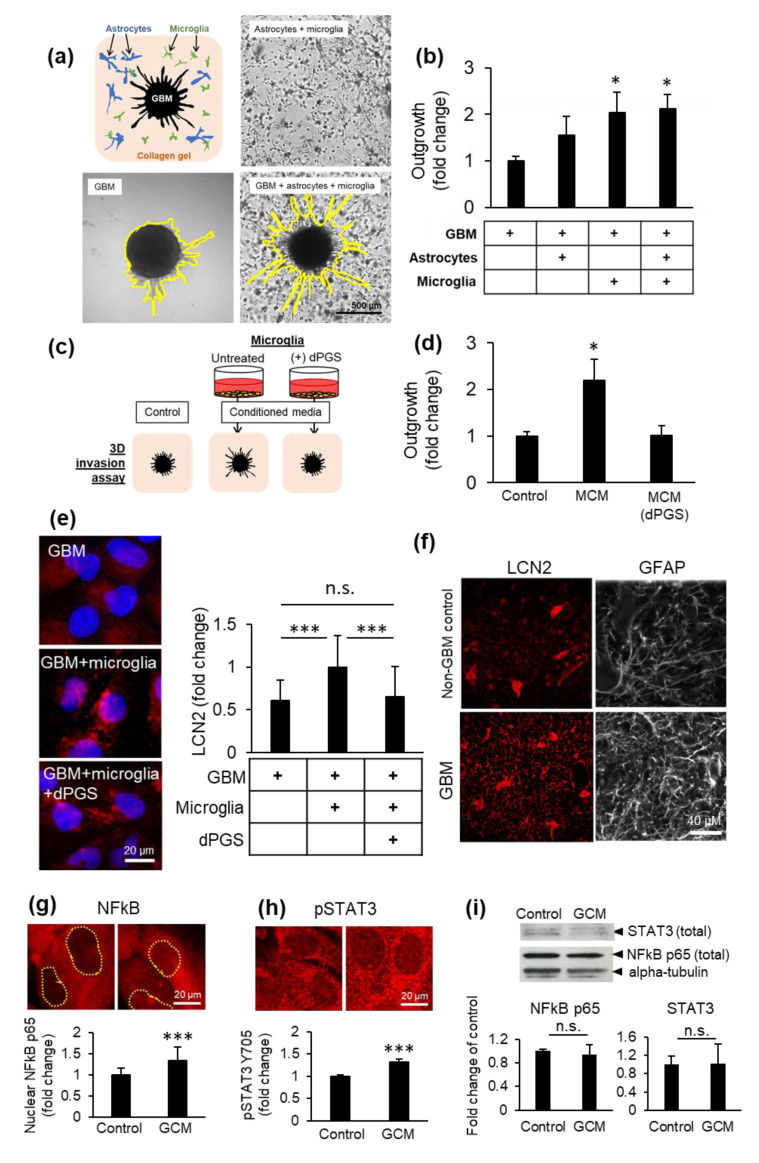
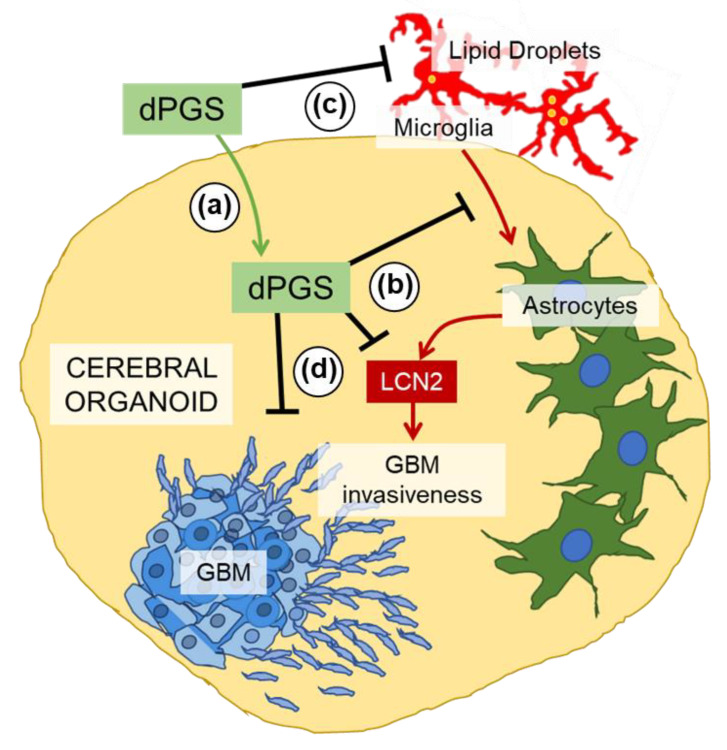
Inflammatory processes in the brain are orchestrated by microglia and astrocytes in response to activators such as pathogen-associated molecular patterns, danger-associated molecular patterns and some nanostructures. Microglia are the primary immune responders in the brain and initiate responses amplified by astrocytes through intercellular signaling. Intercellular communication between neural cells can be studied in cerebral organoids, co-cultures or in vivo. We used human cerebral organoids and glioblastoma co-cultures to study glia modulation by dendritic polyglycerol sulfate (dPGS). dPGS is an extensively studied nanostructure with inherent anti-inflammatory properties. Under inflammatory conditions, lipocalin-2 levels in astrocytes are markedly increased and indirectly enhanced by soluble factors released from hyperactive microglia. dPGS is an effective anti-inflammatory modulator of these markers. Our results show that dPGS can enter neural cells in cerebral organoids and glial cells in monocultures in a time-dependent manner. dPGS markedly reduces lipocalin-2 abundance in the neural cells. Glioblastoma tumoroids of astrocytic origin respond to activated microglia with enhanced invasiveness, whereas conditioned media from dPGS-treated microglia reduce tumoroid invasiveness. Considering that many nanostructures have only been tested in cancer cells and rodent models, experiments in human 3D cerebral organoids and co-cultures are complementary in vitro models to evaluate nanotherapeutics in the pre-clinical setting. Thoroughly characterized organoids and standardized procedures for their preparation are prerequisites to gain information of translational value in nanomedicine. This study provides data for a well-characterized dendrimer (dPGS) that modulates the activation state of human microglia implicated in brain tumor invasiveness.
Keywords: astrocytes; cerebral organoids; glioblastoma; inflammation; lipocalin-2; microglia; nanomedicines.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
