

Identification of Zinc-Dependent Mechanisms Used by Group B Streptococcus To Overcome Calprotectin-Mediated Stress
- PMID: 33173000
- PMCID: PMC7667036
- DOI: 10.1128/mBio.02302-20
Identification of Zinc-Dependent Mechanisms Used by Group B Streptococcus To Overcome Calprotectin-Mediated Stress
Abstract
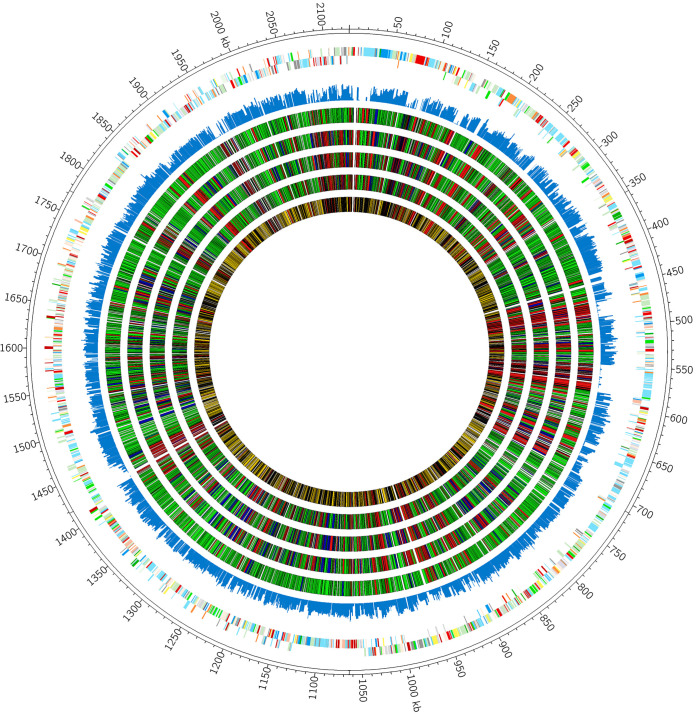
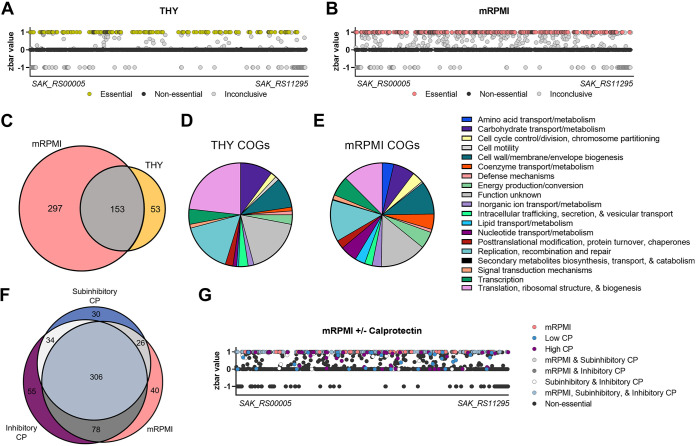
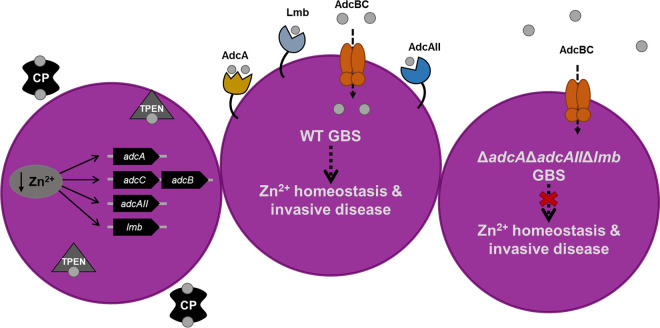
Nutritional immunity is an elegant host mechanism used to starve invading pathogens of necessary nutrient metals. Calprotectin, a metal-binding protein, is produced abundantly by neutrophils and is found in high concentrations within inflammatory sites during infection. Group B Streptococcus (GBS) colonizes the gastrointestinal and female reproductive tracts and is commonly associated with severe invasive infections in newborns such as pneumonia, sepsis, and meningitis. Although GBS infections induce robust neutrophil recruitment and inflammation, the dynamics of GBS and calprotectin interactions remain unknown. Here, we demonstrate that disease and colonizing isolate strains exhibit susceptibility to metal starvation by calprotectin. We constructed a mariner transposon (Krmit) mutant library in GBS and identified 258 genes that contribute to surviving calprotectin stress. Nearly 20% of all underrepresented mutants following treatment with calprotectin are predicted metal transporters, including known zinc systems. As calprotectin binds zinc with picomolar affinity, we investigated the contribution of GBS zinc uptake to overcoming calprotectin-imposed starvation. Quantitative reverse transcriptase PCR (qRT-PCR) revealed a significant upregulation of genes encoding zinc-binding proteins, adcA, adcAII, and lmb, following calprotectin exposure, while growth in calprotectin revealed a significant defect for a global zinc acquisition mutant (ΔadcAΔadcAIIΔlmb) compared to growth of the GBS wild-type (WT) strain. Furthermore, mice challenged with the ΔadcAΔadcAIIΔlmb mutant exhibited decreased mortality and significantly reduced bacterial burden in the brain compared to mice infected with WT GBS; this difference was abrogated in calprotectin knockout mice. Collectively, these data suggest that GBS zinc transport machinery is important for combatting zinc chelation by calprotectin and establishing invasive disease.IMPORTANCE Group B Streptococcus (GBS) asymptomatically colonizes the female reproductive tract but is a common causative agent of meningitis. GBS meningitis is characterized by extensive infiltration of neutrophils carrying high concentrations of calprotectin, a metal chelator. To persist within inflammatory sites and cause invasive disease, GBS must circumvent host starvation attempts. Here, we identified global requirements for GBS survival during calprotectin challenge, including known and putative systems involved in metal ion transport. We characterized the role of zinc import in tolerating calprotectin stress in vitro and in a mouse model of infection. We observed that a global zinc uptake mutant was less virulent than the parental GBS strain and found calprotectin knockout mice to be equally susceptible to infection by wild-type (WT) and mutant strains. These findings suggest that calprotectin production at the site of infection results in a zinc-limited environment and reveals the importance of GBS metal homeostasis to invasive disease.
Keywords: GBS; calprotectin; meningitis; nutritional immunity; zinc.
Copyright © 2020 Burcham et al.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
