

Adaptive venom evolution and toxicity in octopods is driven by extensive novel gene formation, expansion, and loss
- PMID: 33175168
- PMCID: PMC7656900
- DOI: 10.1093/gigascience/giaa120
Adaptive venom evolution and toxicity in octopods is driven by extensive novel gene formation, expansion, and loss
Abstract
Background: Cephalopods represent a rich system for investigating the genetic basis underlying organismal novelties. This diverse group of specialized predators has evolved many adaptations including proteinaceous venom. Of particular interest is the blue-ringed octopus genus (Hapalochlaena), which are the only octopods known to store large quantities of the potent neurotoxin, tetrodotoxin, within their tissues and venom gland.
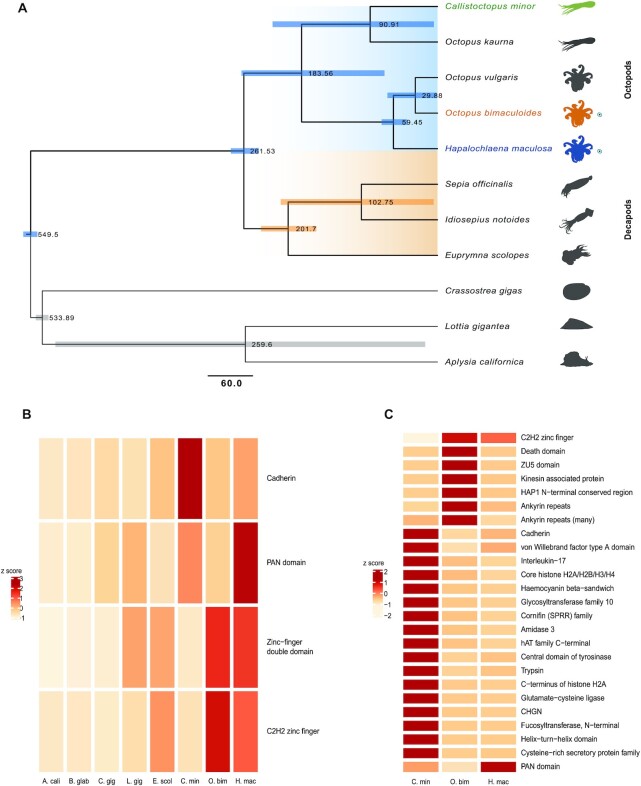
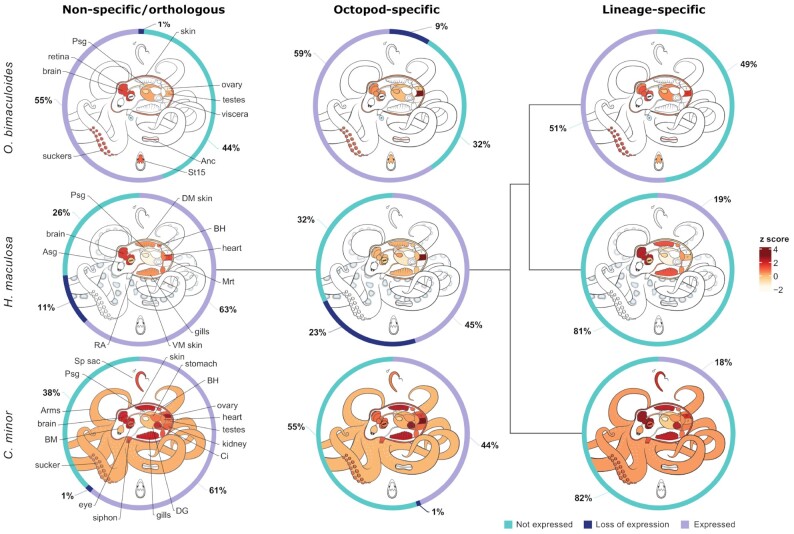
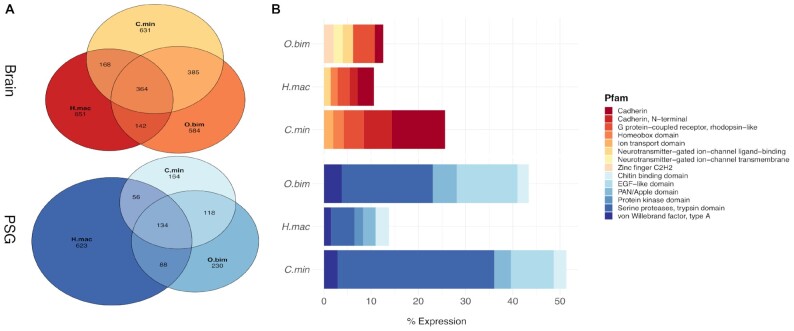
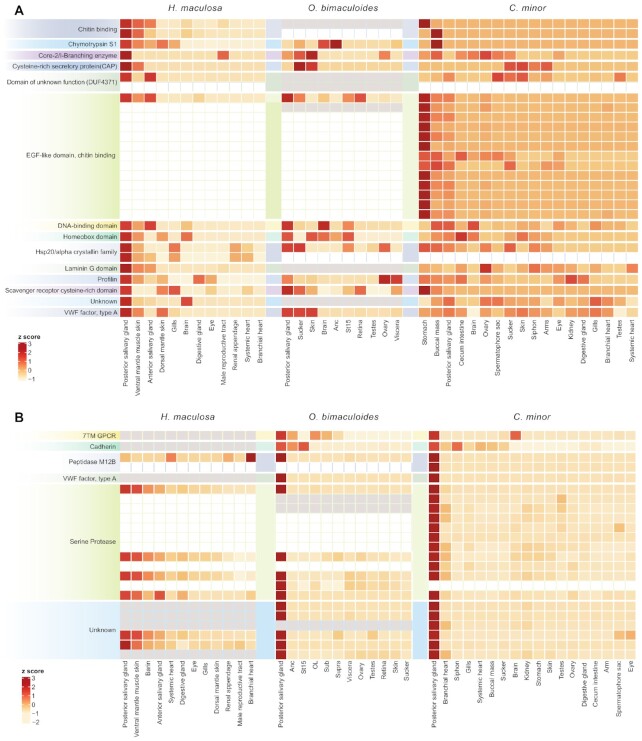
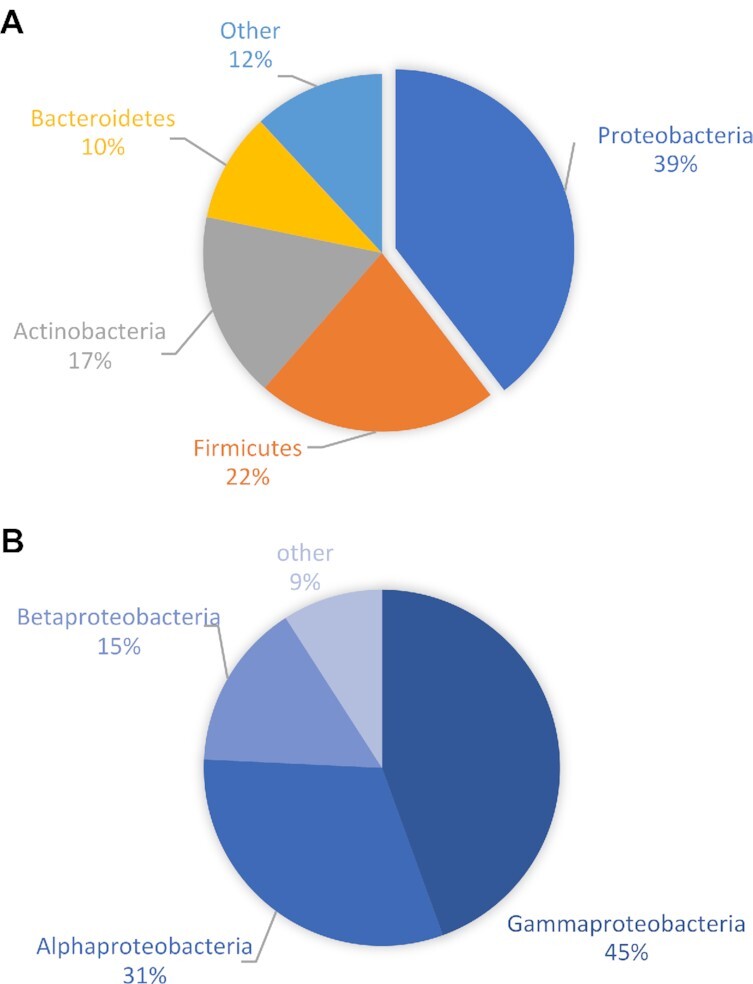
Findings: To reveal genomic correlates of organismal novelties, we conducted a comparative study of 3 octopod genomes, including the Southern blue-ringed octopus (Hapalochlaena maculosa). We present the genome of this species and reveal highly dynamic evolutionary patterns at both non-coding and coding organizational levels. Gene family expansions previously reported in Octopus bimaculoides (e.g., zinc finger and cadherins, both associated with neural functions), as well as formation of novel gene families, dominate the genomic landscape in all octopods. Examination of tissue-specific genes in the posterior salivary gland revealed that expression was dominated by serine proteases in non-tetrodotoxin-bearing octopods, while this family was a minor component in H. maculosa. Moreover, voltage-gated sodium channels in H. maculosa contain a resistance mutation found in pufferfish and garter snakes, which is exclusive to the genus. Analysis of the posterior salivary gland microbiome revealed a diverse array of bacterial species, including genera that can produce tetrodotoxin, suggestive of a possible production source.
Conclusions: We present the first tetrodotoxin-bearing octopod genome H. maculosa, which displays lineage-specific adaptations to tetrodotoxin acquisition. This genome, along with other recently published cephalopod genomes, represents a valuable resource from which future work could advance our understanding of the evolution of genomic novelty in this family.
Keywords: cephalopod genome; comparative genomics; gene family expansions; transposable elements; venom evolution.
© The Author(s) 2020. Published by Oxford University Press GigaScience.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures

References
-
- Ritschard EA, Whitelaw B, Albertin CB, et al. Coupled genomic evolutionary histories as signatures of organismal innovations in cephalopods: Co-evolutionary signatures across levels of genome organization may shed light on functional linkage and origin of cephalopod novelties. Bioessays. 2019;41(12):e1900073. - PubMed
-
- Jereb P, Roper CFE, Norman MD, et al. Cephalopods of the world. An annotated and illustrated catalogue of cephalopod species known to date. In: Octopods and Vampire Squids, Vol. 3. Rome: FAO; 2014.
-
- Graziadei PPC, Gagne HT. Sensory innervation in the rim of the octopus sucker. J Morphol. 1976;150(3):639–79. - PubMed
-
- Froesch D. Projection of chromatophore nerves on the body surface of Octopus vulgaris. Mar Biol. 1973;19(2):153–5.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
