

The Transcription Factor Foxp3 Shapes Regulatory T Cell Identity by Tuning the Activity of trans-Acting Intermediaries
- PMID: 33176163
- PMCID: PMC8363055
- DOI: 10.1016/j.immuni.2020.10.010
The Transcription Factor Foxp3 Shapes Regulatory T Cell Identity by Tuning the Activity of trans-Acting Intermediaries
Abstract
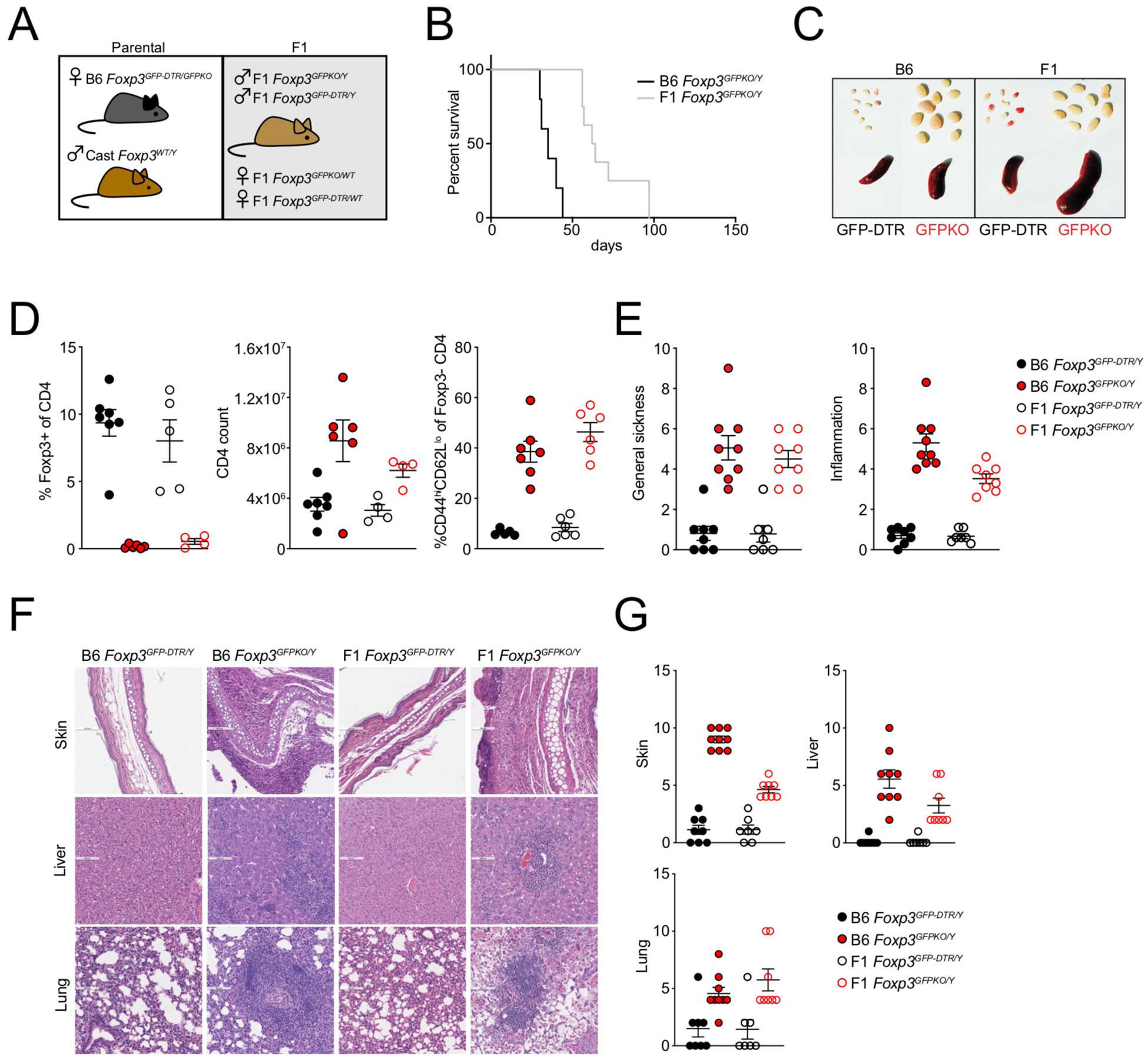
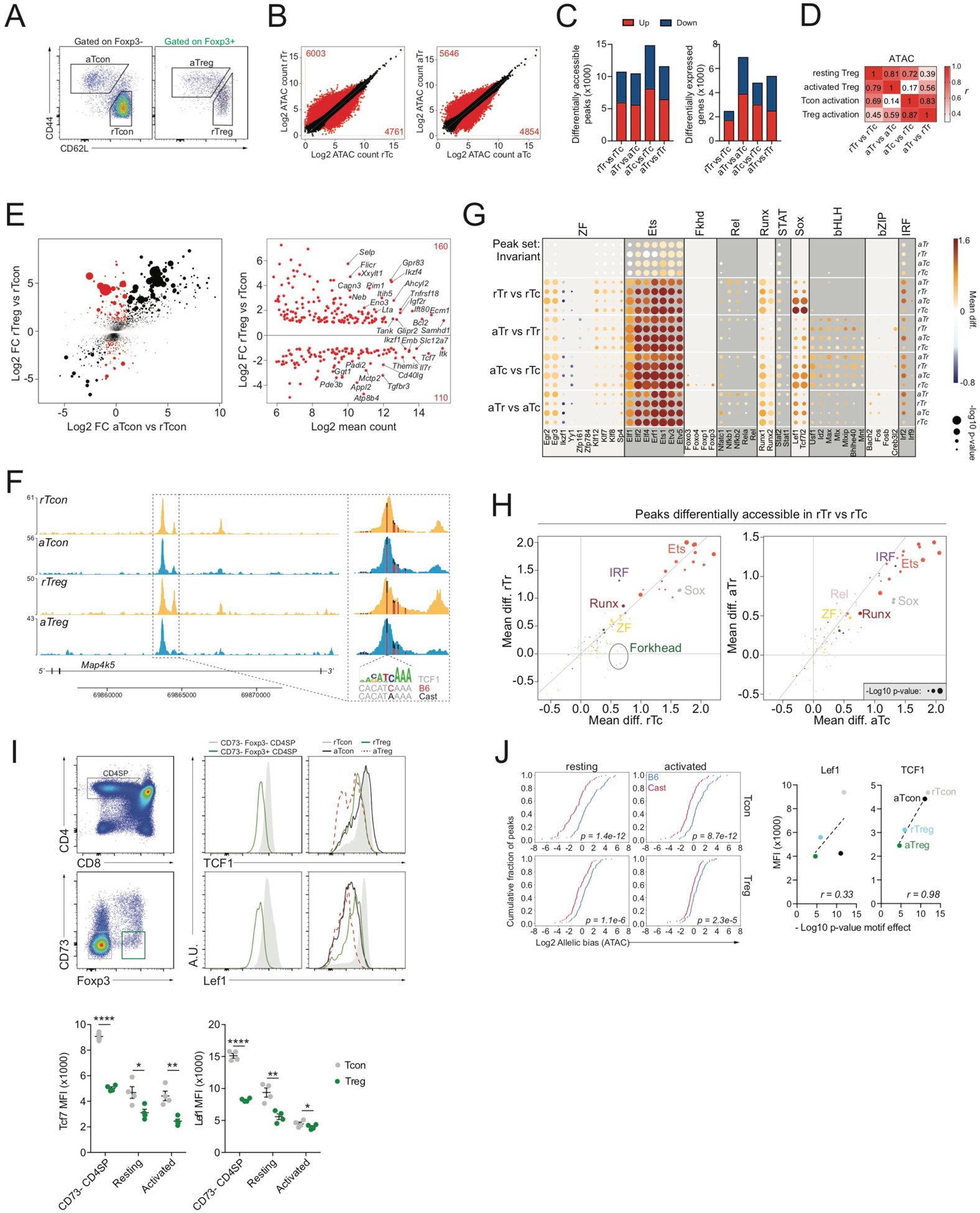
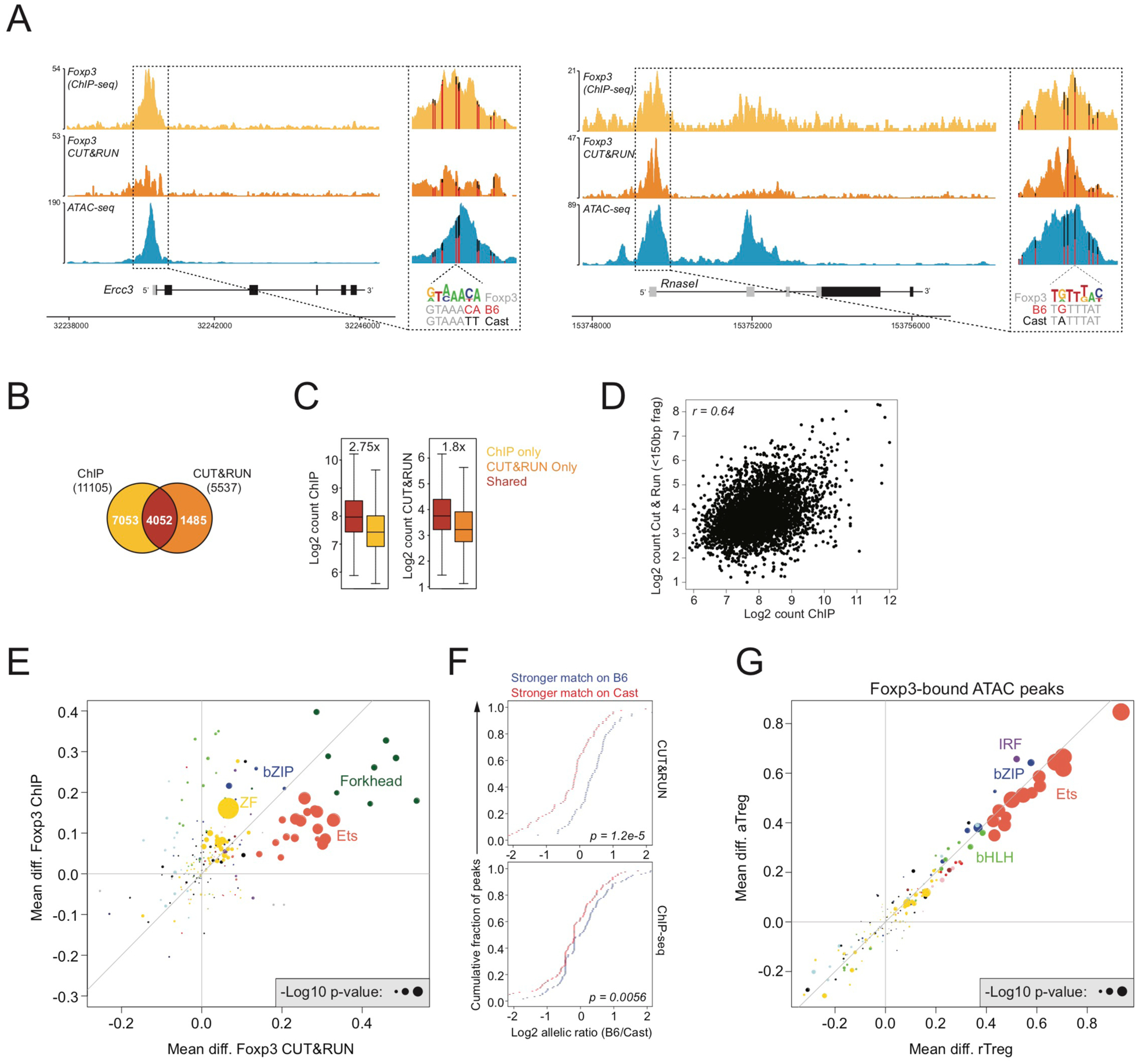
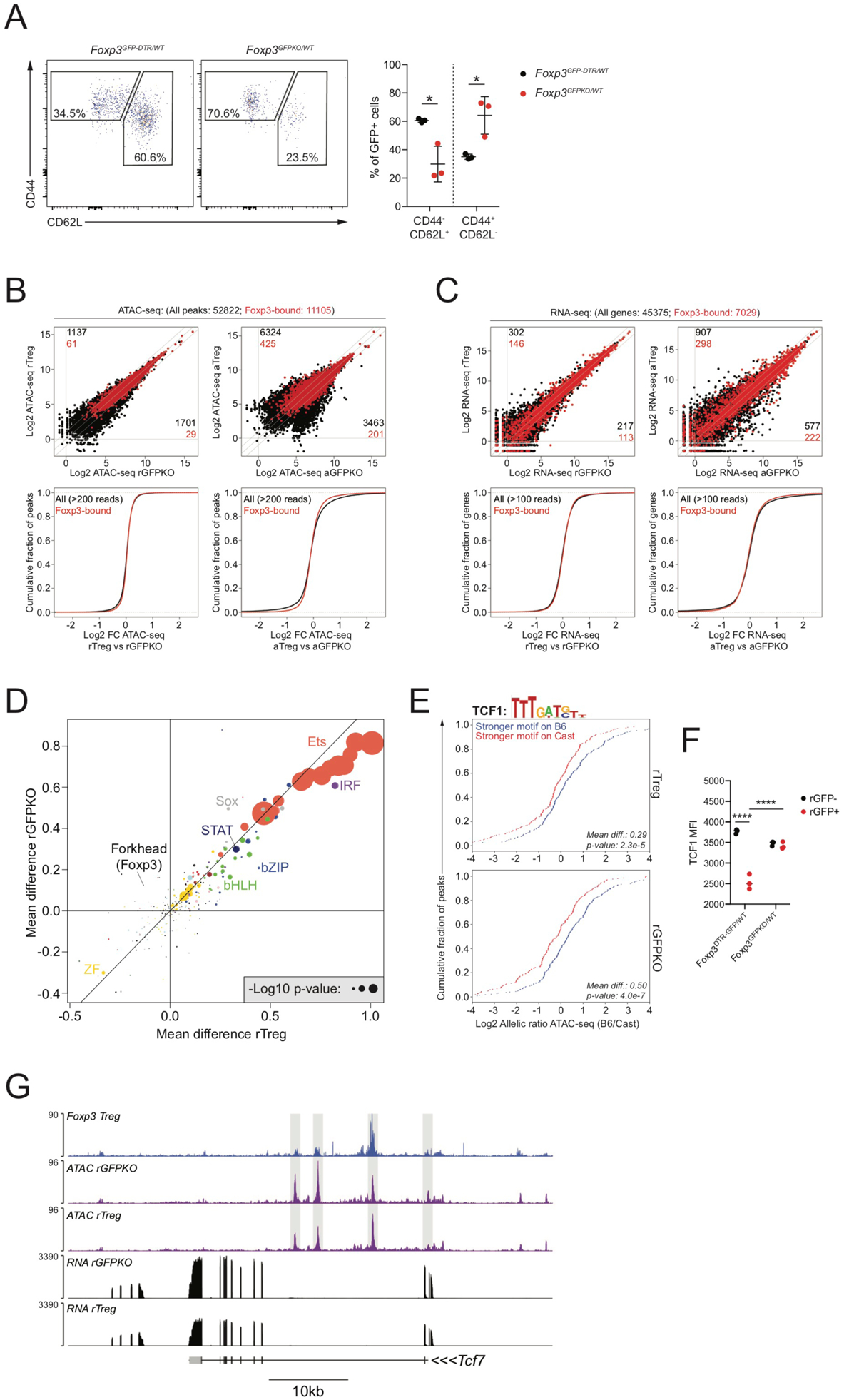
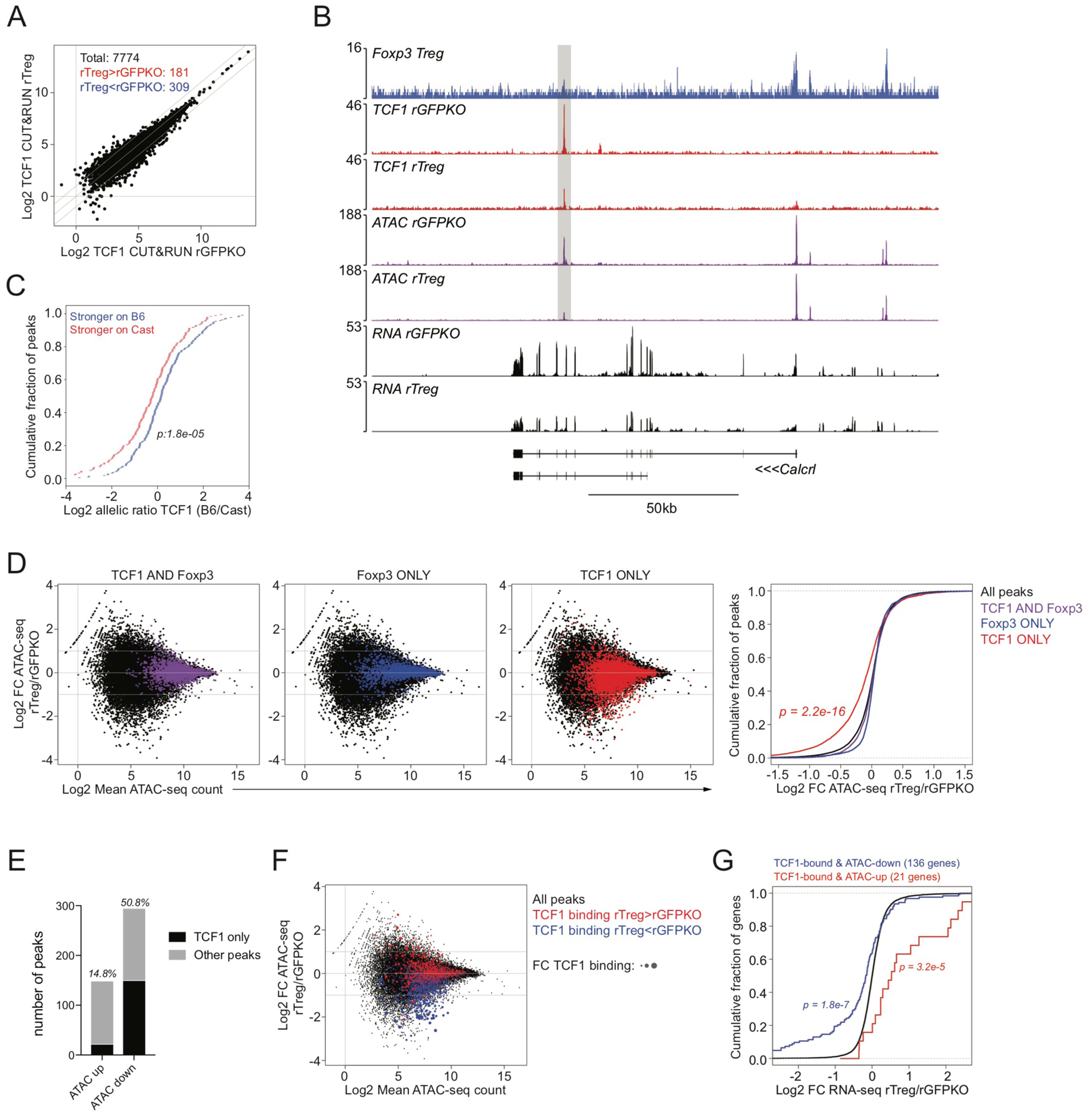
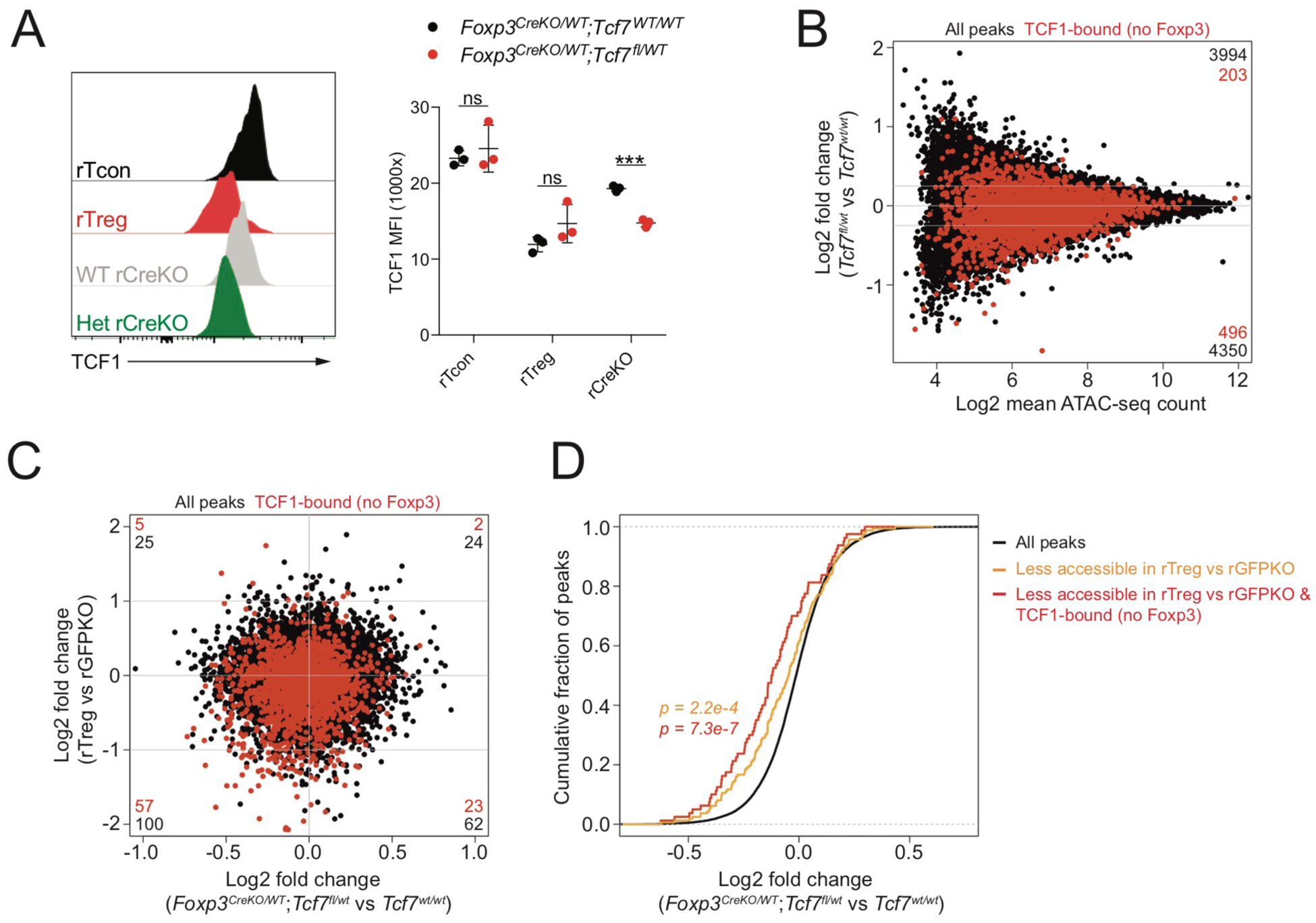
Regulatory T (Treg) cell identity is defined by the lineage-specifying transcription factor (TF) Foxp3. Here we examined mechanisms of Foxp3 function by leveraging naturally occurring genetic variation in wild-derived inbred mice, which enables the identification of DNA sequence motifs driving epigenetic features. Chromatin accessibility, TF binding, and gene expression patterns in resting and activated subsets of Treg cells, conventional CD4 T cells, and cells expressing a Foxp3 reporter null allele revealed that the majority of Foxp3-dependent changes occurred at sites not bound by Foxp3. Chromatin accessibility of these indirect Foxp3 targets depended on the presence of DNA binding motifs for other TFs, including TCF1. Foxp3 expression correlated with decreased TCF1 and reduced accessibility of TCF1-bound chromatin regions. Deleting one copy of the Tcf7 gene recapitulated Foxp3-dependent negative regulation of chromatin accessibility. Thus, Foxp3 defines Treg cell identity in a largely indirect manner by fine-tuning the activity of other major chromatin remodeling TFs such as TCF1.
Copyright © 2020. Published by Elsevier Inc.
Conflict of interest statement
Declaration of Interests A.Y.R. is an SAB member, has equity in Sonoma Biotherapeutics and Vedanta Biosciences, and is a co-inventor or has IP licensed to Takeda that is unrelated to the content of this study.
Figures
Comment in
-
Foxp3 Re-distributes Its Heavy Lifting.Immunity. 2020 Nov 17;53(5):895-897. doi: 10.1016/j.immuni.2020.10.021. Immunity. 2020. PMID: 33207211
References
-
- Bennett CL, Christie J, Ramsdell F, Brunkow ME, Ferguson PJ, Whitesell L, Kelly TE, Saulsbury FT, Chance PF, and Ochs HD (2001). The immune dysregulation, polyendocrinopathy, enteropathy, X-linked syndrome (IPEX) is caused by mutations of FOXP3. Nat Genet 27, 20–21. - PubMed
-
- Boller S, Ramamoorthy S, Akbas D, Nechanitzky R, Burger L, Murr R, Schübeler D, and Grosschedl R (2016). Pioneering Activity of the C-Terminal Domain of EBF1 Shapes the Chromatin Landscape for B Cell Programming. Immunity 44, 527–541. - PubMed
-
- Brunkow ME, Jeffery EW, Hjerrild KA, Paeper B, Clark LB, Yasayko SA, Wilkinson JE, Galas D, Ziegler SF, and Ramsdell F (2001). Disruption of a new forkhead/winged-helix protein, scurfin, results in the fatal lymphoproliferative disorder of the scurfy mouse. Nat Genet 27, 68–73. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
