

Tomatidine inhibits porcine epidemic diarrhea virus replication by targeting 3CL protease
- PMID: 33176871
- PMCID: PMC7656508
- DOI: 10.1186/s13567-020-00865-y
Tomatidine inhibits porcine epidemic diarrhea virus replication by targeting 3CL protease
Abstract
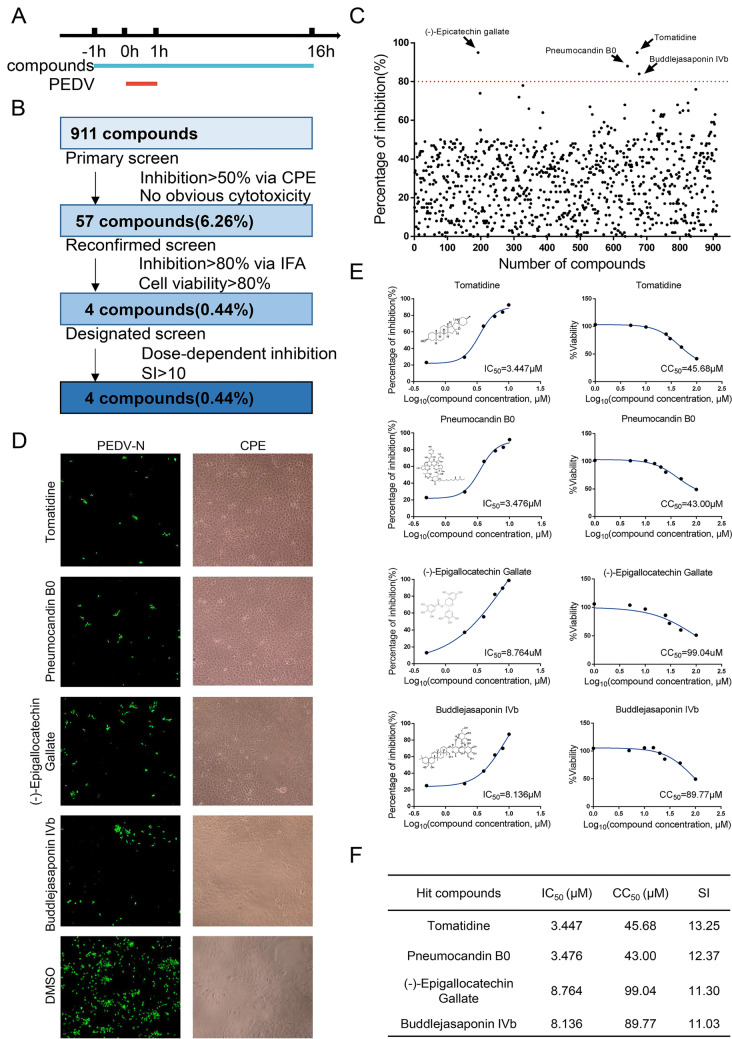
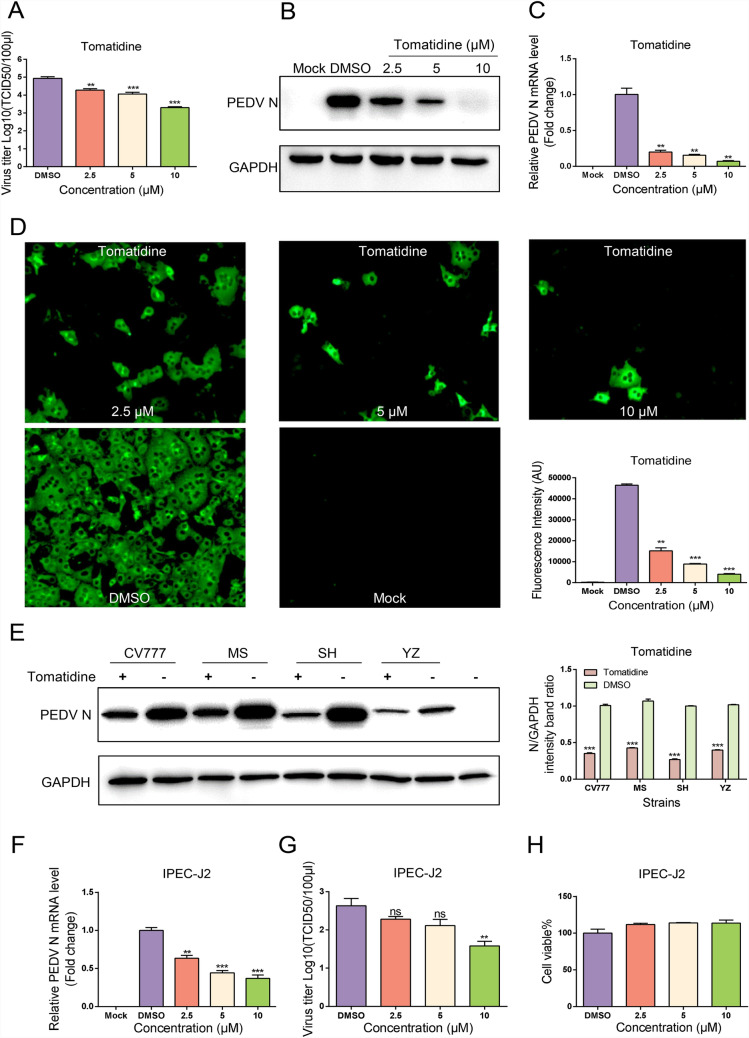
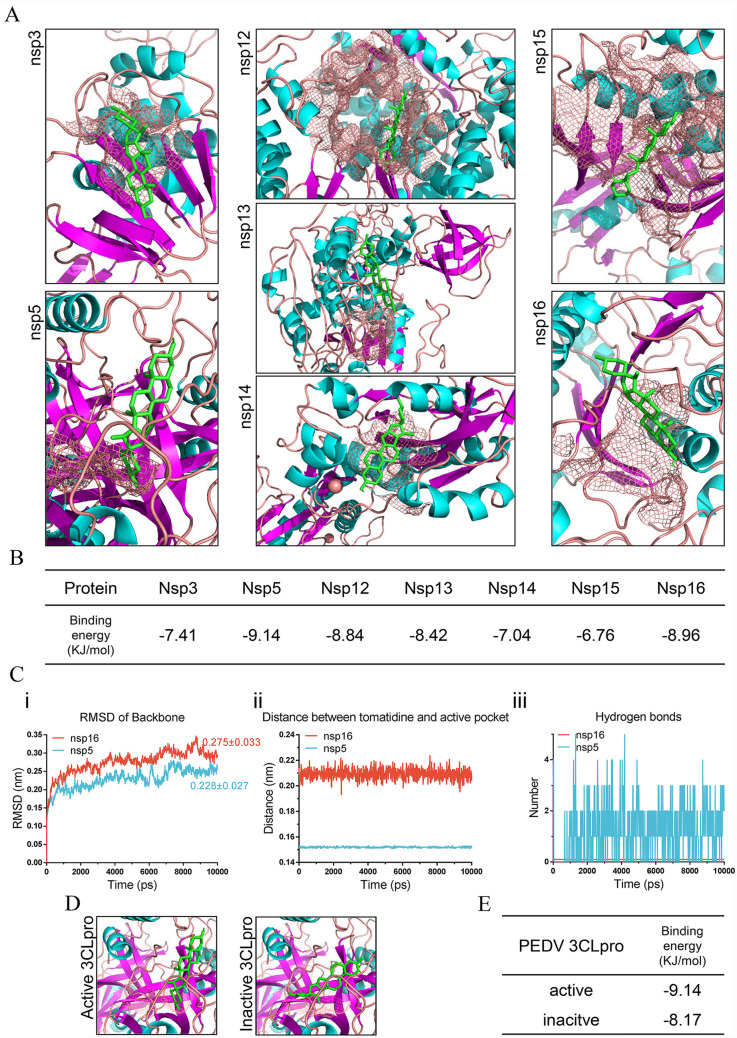
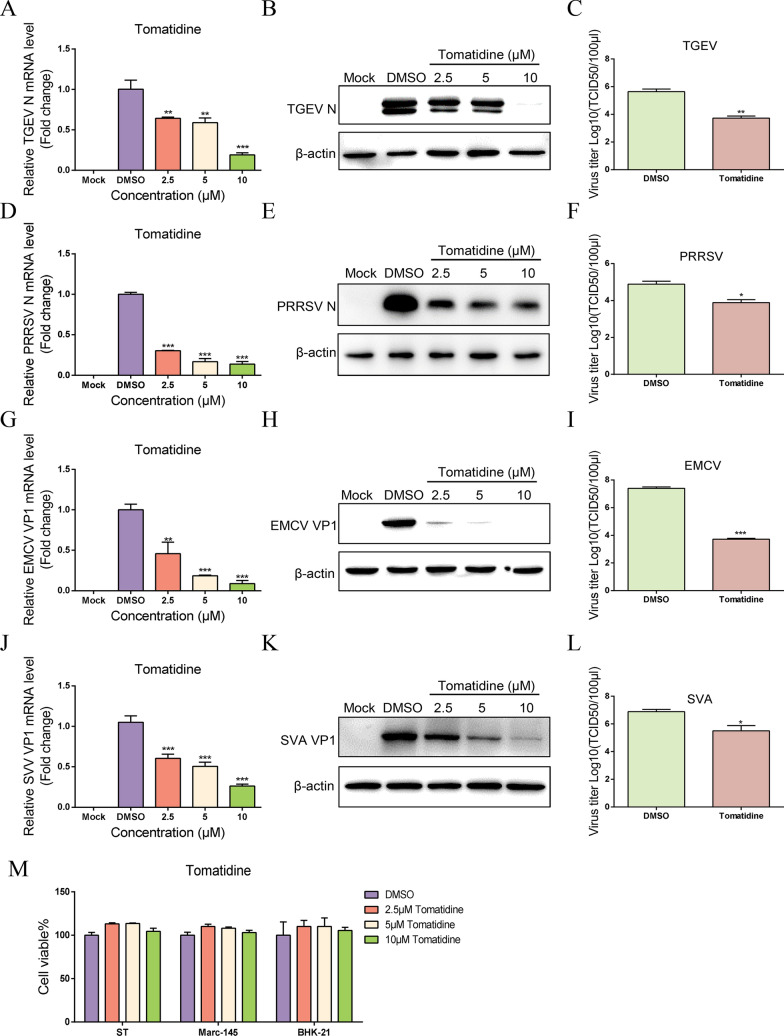
Porcine epidemic diarrhea virus (PEDV) causes lethal diarrhea in suckling piglets, leading to severe economic losses worldwide. There is an urgent need to find new therapeutic methods to prevent and control PEDV. Not only is there a shortage of commercial anti-PEDV drugs, but available commercial vaccines fail to protect against highly virulent PEDV variants. We screened an FDA-approved library of 911 natural products and found that tomatidine, a steroidal alkaloid extracted from the skin and leaves of tomatoes, demonstrates significant inhibition of PEDV replication in Vero and IPEC-J2 cells in vitro. Molecular docking and molecular dynamics analysis predicted interactions between tomatidine and the active pocket of PEDV 3CL protease, which were confirmed by fluorescence spectroscopy and isothermal titration calorimetry (ITC). The inhibiting effect of tomatidine on 3CL protease was determined using cleavage visualization and FRET assay. Tomatidine-mediated blocking of 3CL protease activity in PEDV-infected cells was examined by western blot detection of the viral polyprotein in PEDV-infected cells. It indicates that tomatidine inhibits PEDV replication mainly by targeting 3CL protease. In addition, tomatidine also has antiviral activity against transmissible gastroenteritis virus (TGEV), porcine reproductive and respiratory syndrome virus (PRRSV), encephalo myocarditis virus (EMCV) and seneca virus A (SVA) in vitro. These results may be helpful in developing a new prophylactic and therapeutic strategy against PEDV and other swine disease infections.
Keywords: 3CL protease; PEDV replication; tomatidine.
Conflict of interest statement
None of the authors have any possible conflicts of interest.
Figures



References
-
- Rota PA, Oberste MS, Monroe SS, Nix WA, Campagnoli R, Icenogle JP, Penaranda S, Bankamp B, Maher K, Chen MH, Tong S, Tamin A, Lowe L, Frace M, DeRisi JL, Chen Q, Wang D, Erdman DD, Peret TC, Burns C, Ksiazek TG, Rollin PE, Sanchez A, Liffick S, Holloway B, Limor J, McCaustland K, Olsen-Rasmussen M, Fouchier R, Gunther S, Osterhaus AD, Drosten C, Pallansch MA, Anderson LJ, Bellini WJ. Characterization of a novel coronavirus associated with severe acute respiratory syndrome. Science. 2003;300(5624):1394–1399. doi: 10.1126/science.1085952. - DOI - PubMed
-
- Assiri A, Al-Tawfiq JA, Al-Rabeeah AA, Al-Rabiah FA, Al-Hajjar S, Al-Barrak A, Flemban H, Al-Nassir WN, Balkhy HH, Al-Hakeem RF, Makhdoom HQ, Zumla AI, Memish ZA. Epidemiological, demographic, and clinical characteristics of 47 cases of Middle East respiratory syndrome coronavirus disease from Saudi Arabia: a descriptive study. Lancet Infect Dis. 2013;13(9):752–761. doi: 10.1016/s1473-3099(13)70204-4. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
